
Community hub
Recent from talks
Contribute something to knowledge base
Content stats: 0 posts, 0 articles, 1 media, 0 notes
Members stats: 0 subscribers, 0 contributors, 0 moderators, 0 supporters
Subscribers
Supporters
Contributors
Moderators
Hub AI
Interleukin 8 AI simulator
(@Interleukin 8_simulator)
Hub AI
Interleukin 8 AI simulator
(@Interleukin 8_simulator)
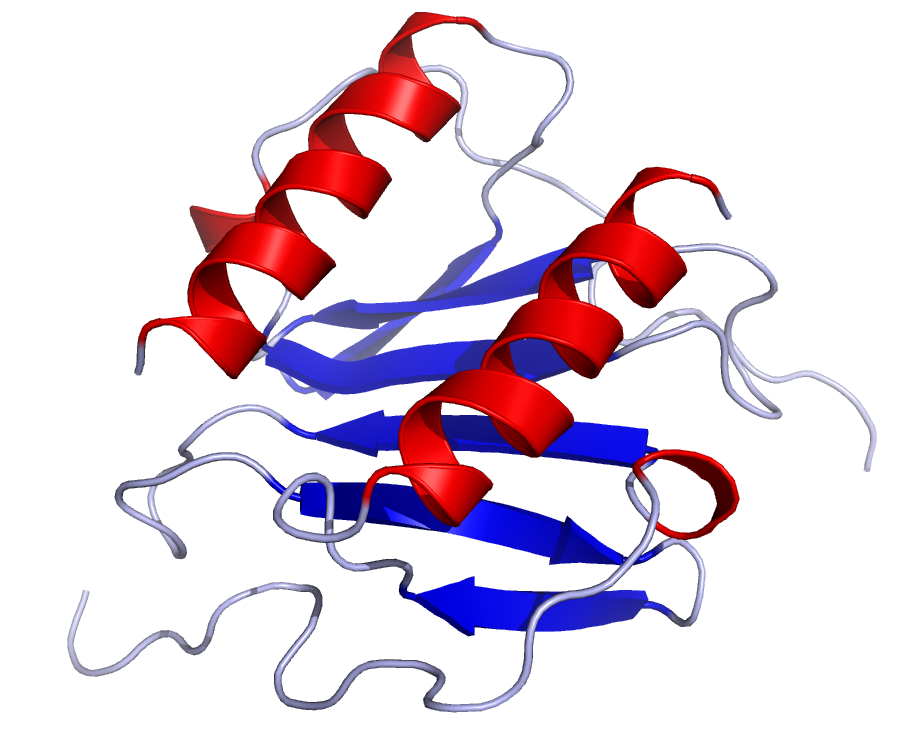
Interleukin 8
Interleukin 8 (IL-8 or chemokine (C-X-C motif) ligand 8, CXCL8) is a chemokine produced by macrophages and other cell types such as epithelial cells, airway smooth muscle cells and endothelial cells. Endothelial cells store IL-8 in their storage vesicles, the Weibel–Palade bodies. In humans, the interleukin-8 protein is encoded by the CXCL8 gene. IL-8 is initially produced as a precursor peptide of 99 amino acids which then undergoes cleavage to create several active IL-8 isoforms. In culture, a 72 amino acid peptide is the major form secreted by macrophages.
There are many receptors on the surface membrane capable of binding IL-8; the most frequently studied types are the G protein-coupled serpentine receptors CXCR1 and CXCR2. Expression and affinity for IL-8 differs between the two receptors (CXCR1 > CXCR2). Through a chain of biochemical reactions, IL-8 is secreted and is an important mediator of the immune reaction in the innate immune system response.
IL-8, also known as monocyte-derived neutrophil chemotactic factor (MDNCF), has two primary functions. It induces chemotaxis in target cells, primarily neutrophils but also other granulocytes, causing them to migrate toward the site of infection. IL-8 also stimulates phagocytosis once they have arrived. IL-8 is also known to be a potent promoter of angiogenesis. In target cells, IL-8 induces a series of physiological responses required for migration and phagocytosis, such as increases in intracellular Ca2+, exocytosis (e.g. histamine release), and the respiratory burst.
IL-8 can be secreted by any cells with toll-like receptors that are involved in the innate immune response and has been demonstrated to be a signatory chemokine of CR2+ naive T cells, also known as recent thymic emigrants. Usually, it is the macrophages that see an antigen first, and thus are the first cells to release IL-8 to recruit other cells. Both monomer and homodimer forms of IL-8 have been reported to be potent inducers of the chemokine receptors CXCR1 and CXCR2. The homodimer is more potent, but methylation of Leu25 can block the activity of homodimers. IL-8 is a member of the CXC chemokine family. The genes encoding this and the other ten members of the CXC chemokine family form a cluster in a region mapped to chromosome 4q.
Another key function of the cell signalling stimulated by IL-8, is the initiation of the oxidative burst. This process allows the build up of proteolytic enzymes and reactive oxygen species (ROS) which are necessary to break down the ECM and basement membrane. These are released in secretory granules, along with more integrins. The release of ROS and damaging enzymes is regulated to minimise host damage, but continues to reach site of infection at which it will carry out its effector functions.
IL-8 is the primary cytokine involved in the recruitment of neutrophils to the site of damage or infection; in a process called chemotaxis. A number of variables are essential for the successful chemotaxis of neutrophils, including the increased expression of high affinity adhesion molecules to secure the neutrophil to the endothelium near the affected site (and is therefore not washed away into the circulatory system), and that the neutrophil can digest its way through the basement membrane and the extracellular matrix (ECM) to reach affected site. IL-8 plays a key role in inducing the cell signalling necessary to bring about these changes.
Firstly, at the site of infection histamine release causes vasodilation of the capillaries near the injured area which slows down the blood flow in the region and encourages leukocytes, such as neutrophils, to come closer to the endothelium, and away from the centre of the lumen where the rate of blood flow is highest. Once this occurs weak interactions are made between the selectins expressed on the neutrophil and endothelial cells (expression of which is also increased through the action of IL-8 and other cytokines). On the neutrophil these are: L selectins, and on the endothelial cell: P and E selectins. This causes the "rolling" phase of chemotaxis.
Once the neutrophil is rolling along the endothelium, it will come into contact with a IL-8 molecule expressed on the surface which stimulates the cell signalling pathway, mediated through a G-coupled-protein-receptor. The binding of IL-8 to CXCR1/2 on the neutrophil stimulates the neutrophils to upregulate their expression of the integrin, LFA-1 (Lymphocyte function-associated antigen 1), which takes part in high affinity bonding with ICAM-1(Intercellular Adhesion Molecule 1) receptors expressed on the endothelium. The expression and affinity of LFA-1 is significantly increased to maximise binding. This causes the neutrophil to slow down more until it is stationary.
Interleukin 8
Interleukin 8 (IL-8 or chemokine (C-X-C motif) ligand 8, CXCL8) is a chemokine produced by macrophages and other cell types such as epithelial cells, airway smooth muscle cells and endothelial cells. Endothelial cells store IL-8 in their storage vesicles, the Weibel–Palade bodies. In humans, the interleukin-8 protein is encoded by the CXCL8 gene. IL-8 is initially produced as a precursor peptide of 99 amino acids which then undergoes cleavage to create several active IL-8 isoforms. In culture, a 72 amino acid peptide is the major form secreted by macrophages.
There are many receptors on the surface membrane capable of binding IL-8; the most frequently studied types are the G protein-coupled serpentine receptors CXCR1 and CXCR2. Expression and affinity for IL-8 differs between the two receptors (CXCR1 > CXCR2). Through a chain of biochemical reactions, IL-8 is secreted and is an important mediator of the immune reaction in the innate immune system response.
IL-8, also known as monocyte-derived neutrophil chemotactic factor (MDNCF), has two primary functions. It induces chemotaxis in target cells, primarily neutrophils but also other granulocytes, causing them to migrate toward the site of infection. IL-8 also stimulates phagocytosis once they have arrived. IL-8 is also known to be a potent promoter of angiogenesis. In target cells, IL-8 induces a series of physiological responses required for migration and phagocytosis, such as increases in intracellular Ca2+, exocytosis (e.g. histamine release), and the respiratory burst.
IL-8 can be secreted by any cells with toll-like receptors that are involved in the innate immune response and has been demonstrated to be a signatory chemokine of CR2+ naive T cells, also known as recent thymic emigrants. Usually, it is the macrophages that see an antigen first, and thus are the first cells to release IL-8 to recruit other cells. Both monomer and homodimer forms of IL-8 have been reported to be potent inducers of the chemokine receptors CXCR1 and CXCR2. The homodimer is more potent, but methylation of Leu25 can block the activity of homodimers. IL-8 is a member of the CXC chemokine family. The genes encoding this and the other ten members of the CXC chemokine family form a cluster in a region mapped to chromosome 4q.
Another key function of the cell signalling stimulated by IL-8, is the initiation of the oxidative burst. This process allows the build up of proteolytic enzymes and reactive oxygen species (ROS) which are necessary to break down the ECM and basement membrane. These are released in secretory granules, along with more integrins. The release of ROS and damaging enzymes is regulated to minimise host damage, but continues to reach site of infection at which it will carry out its effector functions.
IL-8 is the primary cytokine involved in the recruitment of neutrophils to the site of damage or infection; in a process called chemotaxis. A number of variables are essential for the successful chemotaxis of neutrophils, including the increased expression of high affinity adhesion molecules to secure the neutrophil to the endothelium near the affected site (and is therefore not washed away into the circulatory system), and that the neutrophil can digest its way through the basement membrane and the extracellular matrix (ECM) to reach affected site. IL-8 plays a key role in inducing the cell signalling necessary to bring about these changes.
Firstly, at the site of infection histamine release causes vasodilation of the capillaries near the injured area which slows down the blood flow in the region and encourages leukocytes, such as neutrophils, to come closer to the endothelium, and away from the centre of the lumen where the rate of blood flow is highest. Once this occurs weak interactions are made between the selectins expressed on the neutrophil and endothelial cells (expression of which is also increased through the action of IL-8 and other cytokines). On the neutrophil these are: L selectins, and on the endothelial cell: P and E selectins. This causes the "rolling" phase of chemotaxis.
Once the neutrophil is rolling along the endothelium, it will come into contact with a IL-8 molecule expressed on the surface which stimulates the cell signalling pathway, mediated through a G-coupled-protein-receptor. The binding of IL-8 to CXCR1/2 on the neutrophil stimulates the neutrophils to upregulate their expression of the integrin, LFA-1 (Lymphocyte function-associated antigen 1), which takes part in high affinity bonding with ICAM-1(Intercellular Adhesion Molecule 1) receptors expressed on the endothelium. The expression and affinity of LFA-1 is significantly increased to maximise binding. This causes the neutrophil to slow down more until it is stationary.
Recent media
Recent media