
Community hub
Recent from talks
Contribute something to knowledge base
Content stats: 0 posts, 0 articles, 1 media, 0 notes
Members stats: 0 subscribers, 0 contributors, 0 moderators, 0 supporters
Subscribers
Supporters
Contributors
Moderators
Hub AI
Directed evolution AI simulator
(@Directed evolution_simulator)
Hub AI
Directed evolution AI simulator
(@Directed evolution_simulator)
Directed evolution
Directed evolution (DE) is a method used in protein engineering that mimics the process of natural selection to steer proteins or nucleic acids toward a user-defined goal. It consists of subjecting a gene to iterative rounds of mutagenesis (creating a library of variants), selection (expressing those variants and isolating members with the desired function) and amplification (generating a template for the next round). It can be performed in vivo (in living organisms), or in vitro (in cells or free in solution). Directed evolution is used both for protein engineering as an alternative to rationally designing modified proteins, as well as for experimental evolution studies of fundamental evolutionary principles in a controlled, laboratory environment.
Directed evolution has its origins in the 1960s with the evolution of RNA molecules in the "Spiegelman's Monster" experiment. The concept was extended to protein evolution via evolution of bacteria under selection pressures that favoured the evolution of a single gene in its genome.
Early phage display techniques in the 1980s allowed targeting of mutations and selection to a single protein. This enabled selection of enhanced binding proteins, but was not yet compatible with selection for catalytic activity of enzymes. Methods to evolve enzymes were developed in the 1990s and brought the technique to a wider scientific audience. The field rapidly expanded with new methods for making libraries of gene variants and for screening their activity. The development of directed evolution methods was honored in 2018 with the awarding of the Nobel Prize in Chemistry to Frances Arnold for evolution of enzymes, and George Smith and Gregory Winter for phage display.
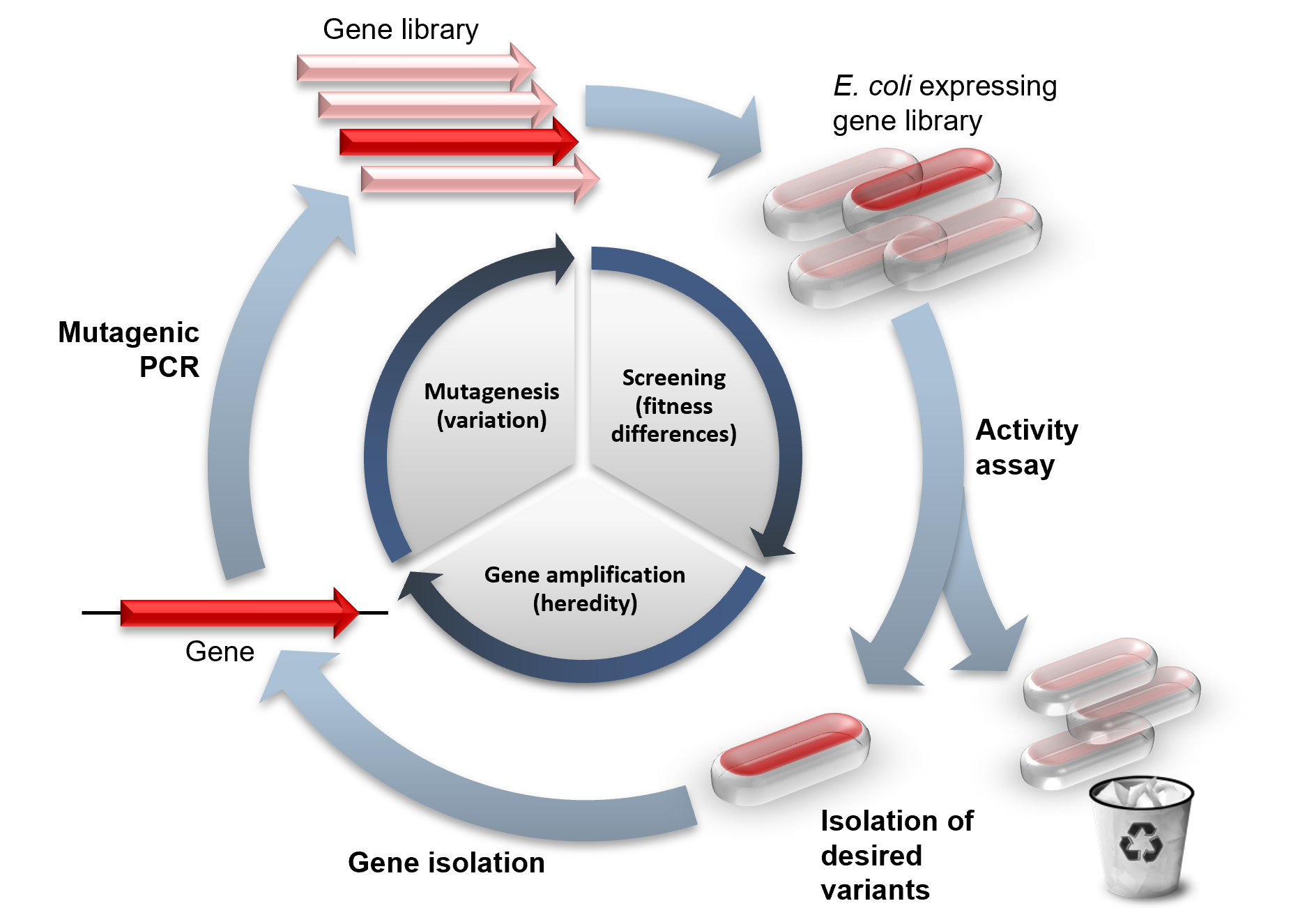
Directed evolution is a mimic of the natural evolution cycle in a laboratory setting. Evolution requires three things to happen: variation between replicators, that the variation causes fitness differences upon which selection acts, and that this variation is heritable. In DE, a single gene is evolved by iterative rounds of mutagenesis, selection or screening, and amplification. Rounds of these steps are typically repeated, using the best variant from one round as the template for the next to achieve stepwise improvements.
The likelihood of success in a directed evolution experiment is directly related to the total library size, as evaluating more mutants increases the chances of finding one with the desired properties.
The first step in performing a cycle of directed evolution is the generation of a library of variant genes. The sequence space for random sequence is vast (10130 possible sequences for a 100 amino acid protein) and extremely sparsely populated by functional proteins. Neither experimental, nor natural[failed verification] evolution can ever get close to sampling so many sequences. Of course, natural evolution samples variant sequences close to functional protein sequences and this is imitated in DE by mutagenising an already functional gene. Some calculations suggest it is entirely feasible that for all practical (i.e. functional and structural) purposes, protein sequence space has been fully explored during the course of evolution of life on Earth.
The starting gene can be mutagenised by random point mutations (by chemical mutagens or error prone PCR) and insertions and deletions (by transposons). Gene recombination can be mimicked by DNA shuffling of several sequences (usually of more than 70% sequence identity) to jump into regions of sequence space between the shuffled parent genes. Finally, specific regions of a gene can be systematically randomised for a more focused approach based on structure and function knowledge. Depending on the method, the library generated will vary in the proportion of functional variants it contains. Even if an organism is used to express the gene of interest, by mutagenising only that gene the rest of the organism's genome remains the same and can be ignored for the evolution experiment (to the extent of providing a constant genetic environment).
The majority of mutations are deleterious and so libraries of mutants tend to mostly have variants with reduced activity. Therefore, a high-throughput assay is vital for measuring activity to find the rare variants with beneficial mutations that improve the desired properties. Two main categories of method exist for isolating functional variants. Selection systems directly couple protein function to survival of the gene, whereas screening systems individually assay each variant and allow a quantitative threshold to be set for sorting a variant or population of variants of a desired activity. Both selection and screening can be performed in living cells (in vivo evolution) or performed directly on the protein or RNA without any cells (in vitro evolution).
Directed evolution
Directed evolution (DE) is a method used in protein engineering that mimics the process of natural selection to steer proteins or nucleic acids toward a user-defined goal. It consists of subjecting a gene to iterative rounds of mutagenesis (creating a library of variants), selection (expressing those variants and isolating members with the desired function) and amplification (generating a template for the next round). It can be performed in vivo (in living organisms), or in vitro (in cells or free in solution). Directed evolution is used both for protein engineering as an alternative to rationally designing modified proteins, as well as for experimental evolution studies of fundamental evolutionary principles in a controlled, laboratory environment.
Directed evolution has its origins in the 1960s with the evolution of RNA molecules in the "Spiegelman's Monster" experiment. The concept was extended to protein evolution via evolution of bacteria under selection pressures that favoured the evolution of a single gene in its genome.
Early phage display techniques in the 1980s allowed targeting of mutations and selection to a single protein. This enabled selection of enhanced binding proteins, but was not yet compatible with selection for catalytic activity of enzymes. Methods to evolve enzymes were developed in the 1990s and brought the technique to a wider scientific audience. The field rapidly expanded with new methods for making libraries of gene variants and for screening their activity. The development of directed evolution methods was honored in 2018 with the awarding of the Nobel Prize in Chemistry to Frances Arnold for evolution of enzymes, and George Smith and Gregory Winter for phage display.
Directed evolution is a mimic of the natural evolution cycle in a laboratory setting. Evolution requires three things to happen: variation between replicators, that the variation causes fitness differences upon which selection acts, and that this variation is heritable. In DE, a single gene is evolved by iterative rounds of mutagenesis, selection or screening, and amplification. Rounds of these steps are typically repeated, using the best variant from one round as the template for the next to achieve stepwise improvements.
The likelihood of success in a directed evolution experiment is directly related to the total library size, as evaluating more mutants increases the chances of finding one with the desired properties.
The first step in performing a cycle of directed evolution is the generation of a library of variant genes. The sequence space for random sequence is vast (10130 possible sequences for a 100 amino acid protein) and extremely sparsely populated by functional proteins. Neither experimental, nor natural[failed verification] evolution can ever get close to sampling so many sequences. Of course, natural evolution samples variant sequences close to functional protein sequences and this is imitated in DE by mutagenising an already functional gene. Some calculations suggest it is entirely feasible that for all practical (i.e. functional and structural) purposes, protein sequence space has been fully explored during the course of evolution of life on Earth.
The starting gene can be mutagenised by random point mutations (by chemical mutagens or error prone PCR) and insertions and deletions (by transposons). Gene recombination can be mimicked by DNA shuffling of several sequences (usually of more than 70% sequence identity) to jump into regions of sequence space between the shuffled parent genes. Finally, specific regions of a gene can be systematically randomised for a more focused approach based on structure and function knowledge. Depending on the method, the library generated will vary in the proportion of functional variants it contains. Even if an organism is used to express the gene of interest, by mutagenising only that gene the rest of the organism's genome remains the same and can be ignored for the evolution experiment (to the extent of providing a constant genetic environment).
The majority of mutations are deleterious and so libraries of mutants tend to mostly have variants with reduced activity. Therefore, a high-throughput assay is vital for measuring activity to find the rare variants with beneficial mutations that improve the desired properties. Two main categories of method exist for isolating functional variants. Selection systems directly couple protein function to survival of the gene, whereas screening systems individually assay each variant and allow a quantitative threshold to be set for sorting a variant or population of variants of a desired activity. Both selection and screening can be performed in living cells (in vivo evolution) or performed directly on the protein or RNA without any cells (in vitro evolution).
Recent media
Recent media