Recent from talks
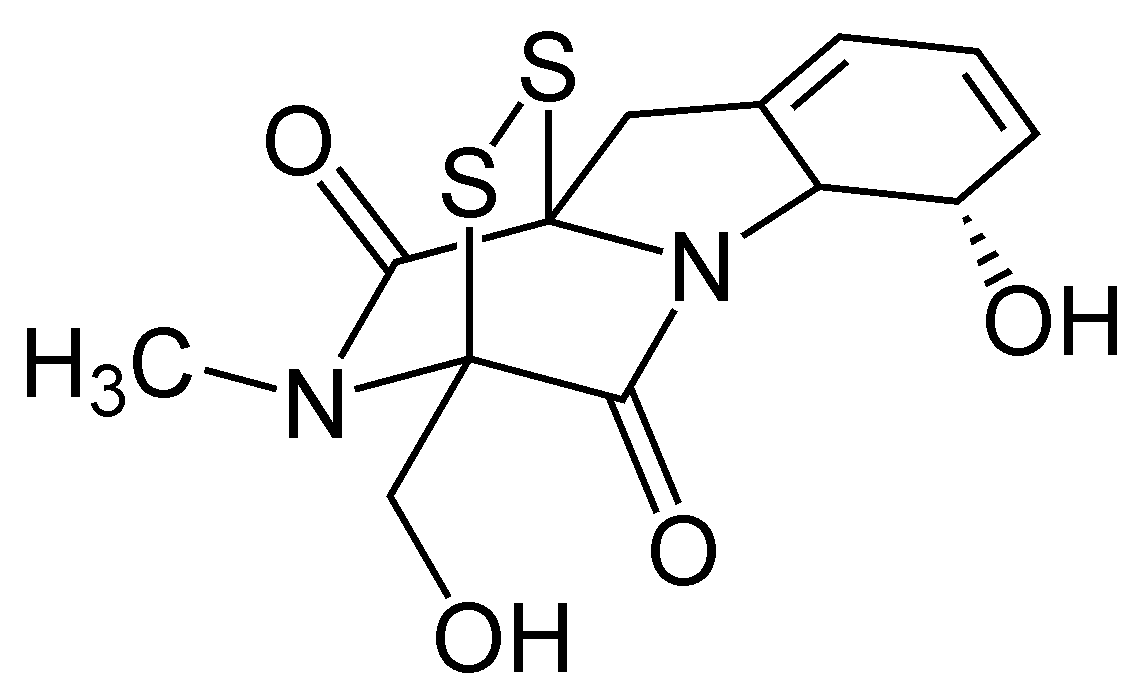
Gliotoxin
Knowledge base stats:
Talk channels stats:
Members stats:
Gliotoxin
Gliotoxin is a sulfur-containing mycotoxin that belongs to a class of naturally occurring 2,5-diketopiperazines produced by several species of fungi, especially those of marine origin. It is the most prominent member of the epipolythiopiperazines, a large class of natural products featuring a diketopiperazine with di- or polysulfide linkage. These highly bioactive compounds have been the subject of numerous studies aimed at new therapeutics. Gliotoxin was originally isolated from Gliocladium fimbriatum, and was named accordingly. It is an epipolythiodioxopiperazine metabolite that is one of the most abundantly produced metabolites in human invasive Aspergillosis (IA).
The compound is produced by human pathogens such as Aspergillus fumigatus, and also by species of Trichoderma and Penicillium. Gliotoxin has also been reported from yeasts of the genus Candida, but results from other studies have cast doubt on the production of this metabolite by Candida fungi. Gliotoxin is not produced by nonpathogenic A. fischeri although A.fischeri contains a gene cluster that is homologous to the gliotoxin gene cluster found in the pathogenic A. fumigatus. Gliotoxin contributes to the pathogenicity of opportunistic fungi by suppressing the immune system response of its host. Gliotoxin additionally possesses fungicidal and bacteriostatic properties, which indicates that it likely plays an important self defense role against bacteria and other fungi for the fungi that produce gliotoxin. Exposure of A. fumigatus to exogenous gliotoxin resulted in aberrant protein expression, especially in those strains that lacked the self-protection protein GliT. There is additional evidence for differential gliotoxin sensitivities amongst fungi including Aspergillus flavus, Fusarium graminearum, and Aspergillus oryzae.
Gliotoxin was first described in 1936 by Weindling and Emerson as a metabolic product from the fungus Trichoderma lignorum. However, afterwards Weindling reported that the fungus had been misidentified based on the advice of C. Thom and M. Timonin, and that the compound instead was isolated from Gliocladium finbriatum. Contention remains on whether the fungus used by Weindling was G. finbriatum or a species of Trichoderma. The chemical structure of gliotoxin was resolved in 1958 by Bell et al. by treatment of gliotoxin on alkaline alumina. Bell and colleagues were able to determine through their structural analyses that the attachment of the disulfide bridge could not occur at any positions other than 3 and 11. This led to the elucidation that gliotoxin was an anhydropeptide related to the amino acids serine and phenylalanine. Additionally, they found that it was noteworthy that the α-carbon atoms of the cooperating α-thio-α-amino acids must have the same configuration.
Gliotoxin is suspected to be an important virulence factor (aka pathogenicity factor) in Aspergillus fungus. Gliotoxin possesses immunosuppressive properties that may suppress and cause apoptosis in certain cells of the immune system, including neutrophils, eosinophils, granulocytes, macrophages, and thymocytes. Specifically, neutrophils exposed to gliotoxin release less reactive oxygen species (ROS) and complete fewer phagocytic activities. Gliotoxin is also believed to interfere with T-cell activation. Additionally, gliotoxin acts as an inhibitor of farnesyl transferase. It noncompetitively inhibits the chymotrypsin-like activity of the 20S proteasome.
In vivo gliotoxin displays anti-inflammatory activity. It was investigated as an antibiotic and antifungal in the 1940s and as an antiviral agent. Gliotoxin inactivates many different enzymes, including nuclear factor-κB (NF-κB), NADPH oxidase, and glutaredoxin. The inhibition of NF-κB leads prevents cytokine release and induction of the inflammatory response.
The immunosuppressive properties of gliotoxin are due to the disulfide bridge within its structure. Interactions occur between sulfur molecules that make up the disulfide bridge and thiol groups contained in cysteine residues. Gliotoxin acts by blocking thiol residues in the cell membrane. Gliotoxin also activates a member of the Bcl-2 family called Bak in order to mediate cell apoptosis. Activated Bak then causes the release of ROS, which form pores within the mitochondrial membrane. These pores allow the release of cytochrome C and AIF, which initiate apoptosis within the cell.
In Aspergillus fumigatus, the enzymes needed for gliotoxin biosynthesis are encoded in 13 genes within the gli gene cluster. When this gene cluster is activated, these enzymes mediate the production of gliotoxin from serine and phenylalanine residues. The function of some genes contained within the gli gene cluster remain to be elucidated.
Enzymes Involved in Biosynthesis (in order of activity)
Hub AI
Gliotoxin AI simulator
(@Gliotoxin_simulator)
Gliotoxin
Gliotoxin is a sulfur-containing mycotoxin that belongs to a class of naturally occurring 2,5-diketopiperazines produced by several species of fungi, especially those of marine origin. It is the most prominent member of the epipolythiopiperazines, a large class of natural products featuring a diketopiperazine with di- or polysulfide linkage. These highly bioactive compounds have been the subject of numerous studies aimed at new therapeutics. Gliotoxin was originally isolated from Gliocladium fimbriatum, and was named accordingly. It is an epipolythiodioxopiperazine metabolite that is one of the most abundantly produced metabolites in human invasive Aspergillosis (IA).
The compound is produced by human pathogens such as Aspergillus fumigatus, and also by species of Trichoderma and Penicillium. Gliotoxin has also been reported from yeasts of the genus Candida, but results from other studies have cast doubt on the production of this metabolite by Candida fungi. Gliotoxin is not produced by nonpathogenic A. fischeri although A.fischeri contains a gene cluster that is homologous to the gliotoxin gene cluster found in the pathogenic A. fumigatus. Gliotoxin contributes to the pathogenicity of opportunistic fungi by suppressing the immune system response of its host. Gliotoxin additionally possesses fungicidal and bacteriostatic properties, which indicates that it likely plays an important self defense role against bacteria and other fungi for the fungi that produce gliotoxin. Exposure of A. fumigatus to exogenous gliotoxin resulted in aberrant protein expression, especially in those strains that lacked the self-protection protein GliT. There is additional evidence for differential gliotoxin sensitivities amongst fungi including Aspergillus flavus, Fusarium graminearum, and Aspergillus oryzae.
Gliotoxin was first described in 1936 by Weindling and Emerson as a metabolic product from the fungus Trichoderma lignorum. However, afterwards Weindling reported that the fungus had been misidentified based on the advice of C. Thom and M. Timonin, and that the compound instead was isolated from Gliocladium finbriatum. Contention remains on whether the fungus used by Weindling was G. finbriatum or a species of Trichoderma. The chemical structure of gliotoxin was resolved in 1958 by Bell et al. by treatment of gliotoxin on alkaline alumina. Bell and colleagues were able to determine through their structural analyses that the attachment of the disulfide bridge could not occur at any positions other than 3 and 11. This led to the elucidation that gliotoxin was an anhydropeptide related to the amino acids serine and phenylalanine. Additionally, they found that it was noteworthy that the α-carbon atoms of the cooperating α-thio-α-amino acids must have the same configuration.
Gliotoxin is suspected to be an important virulence factor (aka pathogenicity factor) in Aspergillus fungus. Gliotoxin possesses immunosuppressive properties that may suppress and cause apoptosis in certain cells of the immune system, including neutrophils, eosinophils, granulocytes, macrophages, and thymocytes. Specifically, neutrophils exposed to gliotoxin release less reactive oxygen species (ROS) and complete fewer phagocytic activities. Gliotoxin is also believed to interfere with T-cell activation. Additionally, gliotoxin acts as an inhibitor of farnesyl transferase. It noncompetitively inhibits the chymotrypsin-like activity of the 20S proteasome.
In vivo gliotoxin displays anti-inflammatory activity. It was investigated as an antibiotic and antifungal in the 1940s and as an antiviral agent. Gliotoxin inactivates many different enzymes, including nuclear factor-κB (NF-κB), NADPH oxidase, and glutaredoxin. The inhibition of NF-κB leads prevents cytokine release and induction of the inflammatory response.
The immunosuppressive properties of gliotoxin are due to the disulfide bridge within its structure. Interactions occur between sulfur molecules that make up the disulfide bridge and thiol groups contained in cysteine residues. Gliotoxin acts by blocking thiol residues in the cell membrane. Gliotoxin also activates a member of the Bcl-2 family called Bak in order to mediate cell apoptosis. Activated Bak then causes the release of ROS, which form pores within the mitochondrial membrane. These pores allow the release of cytochrome C and AIF, which initiate apoptosis within the cell.
In Aspergillus fumigatus, the enzymes needed for gliotoxin biosynthesis are encoded in 13 genes within the gli gene cluster. When this gene cluster is activated, these enzymes mediate the production of gliotoxin from serine and phenylalanine residues. The function of some genes contained within the gli gene cluster remain to be elucidated.
Enzymes Involved in Biosynthesis (in order of activity)
Recent media