
Community hub
Recent from talks
Contribute something to knowledge base
Content stats: 0 posts, 0 articles, 1 media, 0 notes
Members stats: 0 subscribers, 0 contributors, 0 moderators, 0 supporters
Subscribers
Supporters
Contributors
Moderators
Hub AI
KRAS AI simulator
(@KRAS_simulator)
Hub AI
KRAS AI simulator
(@KRAS_simulator)
KRAS
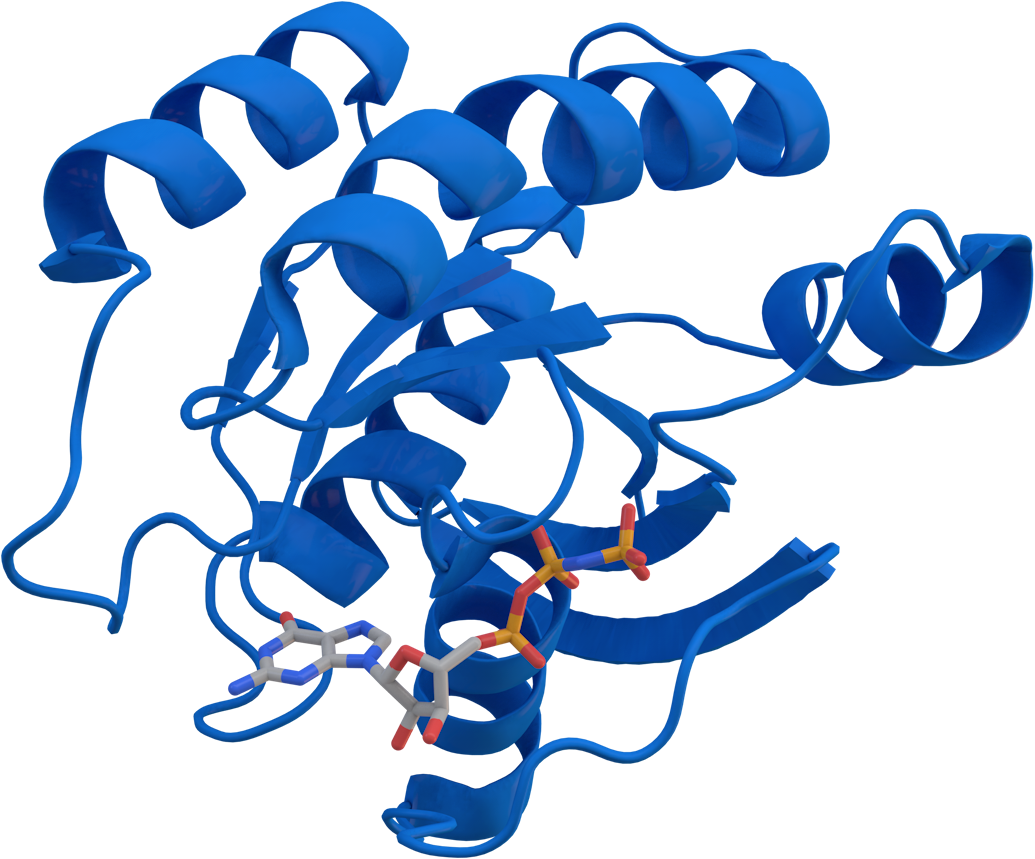
KRAS (Kirsten rat sarcoma virus) is a gene that provides instructions for making a protein called K-Ras, a part of the RAS/MAPK pathway. The protein relays signals from outside the cell to the cell's nucleus. These signals instruct the cell to grow and divide (proliferate) or to mature and take on specialized functions (differentiate). It is called KRAS because it was first identified as a viral oncogene in the Kirsten RAt Sarcoma virus. The oncogene identified was derived from a cellular genome, so KRAS, when found in a cellular genome, is called a proto-oncogene.
The K-Ras protein is a GTPase, a class of enzymes which convert the nucleotide guanosine triphosphate (GTP) into guanosine diphosphate (GDP). In this way the K-Ras protein acts like a switch that is turned on and off by the GTP and GDP molecules. To transmit signals, it must be turned on by attaching (binding) to a molecule of GTP. The K-Ras protein is turned off (inactivated) when it converts the GTP to GDP. When the protein is bound to GDP, it does not relay signals to the nucleus.
The gene product of KRAS, the K-Ras protein, was first found as a p21 GTPase. Like other members of the ras subfamily of GTPases, the K-Ras protein is an early player in many signal transduction pathways. K-Ras is usually tethered to cell membranes because of the farnesylation of its C-terminus. There are two protein products of the KRAS gene in mammalian cells that result from the use of alternative exon 4 (exon 4A and 4B respectively): K-Ras4A and K-Ras4B. These proteins have different structures in their C-terminal region and use different mechanisms to localize to cellular membranes, including the plasma membrane.
KRAS acts as a molecular on/off switch, using protein dynamics. Once it is allosterically activated, it recruits and activates proteins necessary for the propagation of growth factors, as well as other cell signaling receptors like c-Raf and PI 3-kinase. KRAS upregulates the GLUT1 glucose transporter, thereby contributing to the Warburg effect in cancer cells. KRAS binds to GTP in its active state. It also possesses an intrinsic enzymatic activity which cleaves the terminal phosphate of the nucleotide, converting it to GDP. Upon conversion of GTP to GDP, KRAS is deactivated. The rate of conversion is usually slow, but can be increased dramatically by an accessory protein of the GTPase-activating protein (GAP) class, for example RasGAP.< In turn, KRAS can bind to proteins of the Guanine Nucleotide Exchange Factor (GEF) class (such as SOS1), which forces the release of bound nucleotide (GDP). Subsequently, KRAS binds GTP present in the cytosol and the GEF is released from ras-GTP.
Other members of the Ras family include: HRAS and NRAS. These proteins all are regulated in the same manner and appear to differ in their sites of action within the cell.
This proto-oncogene is a Kirsten ras oncogene homolog from the mammalian Ras gene family. A single amino acid substitution, and in particular a single nucleotide substitution, is responsible for an activating mutation. The transforming protein that results is implicated in various malignancies, including lung adenocarcinoma, mucinous adenoma, ductal carcinoma of the pancreas and colorectal cancer.
Several germline KRAS mutations have been found to be associated with Noonan syndrome and cardio-facio-cutaneous syndrome. Somatic KRAS mutations are found at high rates in leukemias, colorectal cancer, pancreatic cancer and lung cancer.
The impact of KRAS mutations is heavily dependent on the order of mutations. Primary KRAS mutations generally lead to a self-limiting hyperplastic or borderline lesion, but if they occur after a previous APC mutation it often progresses to cancer. KRAS mutations are more commonly observed in cecal cancers than colorectal cancers located in any other places from ascending colon to rectum.
KRAS
KRAS (Kirsten rat sarcoma virus) is a gene that provides instructions for making a protein called K-Ras, a part of the RAS/MAPK pathway. The protein relays signals from outside the cell to the cell's nucleus. These signals instruct the cell to grow and divide (proliferate) or to mature and take on specialized functions (differentiate). It is called KRAS because it was first identified as a viral oncogene in the Kirsten RAt Sarcoma virus. The oncogene identified was derived from a cellular genome, so KRAS, when found in a cellular genome, is called a proto-oncogene.
The K-Ras protein is a GTPase, a class of enzymes which convert the nucleotide guanosine triphosphate (GTP) into guanosine diphosphate (GDP). In this way the K-Ras protein acts like a switch that is turned on and off by the GTP and GDP molecules. To transmit signals, it must be turned on by attaching (binding) to a molecule of GTP. The K-Ras protein is turned off (inactivated) when it converts the GTP to GDP. When the protein is bound to GDP, it does not relay signals to the nucleus.
The gene product of KRAS, the K-Ras protein, was first found as a p21 GTPase. Like other members of the ras subfamily of GTPases, the K-Ras protein is an early player in many signal transduction pathways. K-Ras is usually tethered to cell membranes because of the farnesylation of its C-terminus. There are two protein products of the KRAS gene in mammalian cells that result from the use of alternative exon 4 (exon 4A and 4B respectively): K-Ras4A and K-Ras4B. These proteins have different structures in their C-terminal region and use different mechanisms to localize to cellular membranes, including the plasma membrane.
KRAS acts as a molecular on/off switch, using protein dynamics. Once it is allosterically activated, it recruits and activates proteins necessary for the propagation of growth factors, as well as other cell signaling receptors like c-Raf and PI 3-kinase. KRAS upregulates the GLUT1 glucose transporter, thereby contributing to the Warburg effect in cancer cells. KRAS binds to GTP in its active state. It also possesses an intrinsic enzymatic activity which cleaves the terminal phosphate of the nucleotide, converting it to GDP. Upon conversion of GTP to GDP, KRAS is deactivated. The rate of conversion is usually slow, but can be increased dramatically by an accessory protein of the GTPase-activating protein (GAP) class, for example RasGAP.< In turn, KRAS can bind to proteins of the Guanine Nucleotide Exchange Factor (GEF) class (such as SOS1), which forces the release of bound nucleotide (GDP). Subsequently, KRAS binds GTP present in the cytosol and the GEF is released from ras-GTP.
Other members of the Ras family include: HRAS and NRAS. These proteins all are regulated in the same manner and appear to differ in their sites of action within the cell.
This proto-oncogene is a Kirsten ras oncogene homolog from the mammalian Ras gene family. A single amino acid substitution, and in particular a single nucleotide substitution, is responsible for an activating mutation. The transforming protein that results is implicated in various malignancies, including lung adenocarcinoma, mucinous adenoma, ductal carcinoma of the pancreas and colorectal cancer.
Several germline KRAS mutations have been found to be associated with Noonan syndrome and cardio-facio-cutaneous syndrome. Somatic KRAS mutations are found at high rates in leukemias, colorectal cancer, pancreatic cancer and lung cancer.
The impact of KRAS mutations is heavily dependent on the order of mutations. Primary KRAS mutations generally lead to a self-limiting hyperplastic or borderline lesion, but if they occur after a previous APC mutation it often progresses to cancer. KRAS mutations are more commonly observed in cecal cancers than colorectal cancers located in any other places from ascending colon to rectum.
Recent media
Recent media