Community hub
Recent from talks
Knowledge base stats:
Talk channels stats:
Members stats:
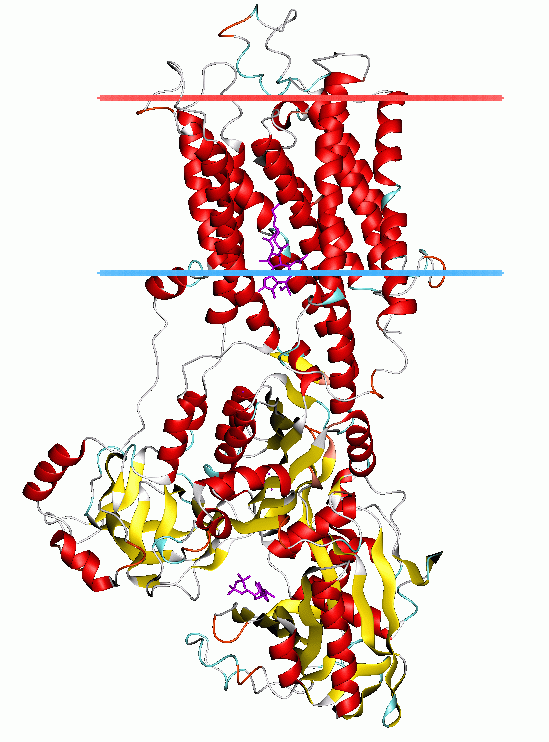
P-type ATPase
The P-type ATPases, also known as E1-E2 ATPases, are a large group of evolutionarily related ion and lipid pumps that are found in bacteria, archaea, and eukaryotes. P-type ATPases are α-helical bundle primary transporters named based upon their ability to catalyze auto- (or self-) phosphorylation (hence P) of a key conserved aspartate residue within the pump and their energy source, adenosine triphosphate (ATP). In addition, they all appear to interconvert between at least two different conformations, denoted by E1 and E2. P-type ATPases fall under the P-type ATPase (P-ATPase) Superfamily (TC# 3.A.3) which, as of early 2016, includes 20 different protein families.
Most members of this transporter superfamily drive cation uptake or efflux, though one subfamily, the flippases, is involved in flipping phospholipids to maintain the asymmetric nature of the biomembrane.
In humans, P-type ATPases serve as a basis for nerve impulses, relaxation of muscles, secretion and absorption in the kidney, absorption of nutrient in the intestine and other physiological processes. Prominent examples of P-type ATPases are the sodium-potassium pump (Na+/K+-ATPase), the proton-potassium pump (H+/K+-ATPase), the calcium pump (Ca2+-ATPase) and the plasma membrane proton pump (H+-ATPase) of plants and fungi.
The generalized reaction for P-type ATPases is
nLigand1 (out) + mLigand2 (in) + ATP → nLigand1 (in) + mLigand2 (out) + ADP + Pi.
where the ligand can be either a metal ion or a phospholipid molecule.
The first P-type ATPase discovered was the Na+/K+-ATPase, which Nobel laureate Jens Christian Skou isolated in 1957. The Na+/K+-ATPase was only the first member of a large and still-growing protein family (see Swiss-Prot Prosite motif PS00154).
P-type ATPases have a single catalytic subunit of 70 - 140 kDa. The catalytic subunit hydrolyzes ATP, contains the aspartyl phosphorylation site and binding sites for the transported ligand(s) and catalyzes ion transport. Various subfamilies of P-type ATPases also need additional subunits for proper function. Additional subunits that lack catalytic activity are present in the ATPase complexes of P1A, P2A, P2C and P4 ATPases. E.g. the catalytic alpha subunit of Na+/K+-ATPase consists of two additional subunits, beta and gamma, involved in trafficking, folding, and regulation of these pumps. The first P-type ATPase to be crystallized was SERCA1a, a sarco(endo)plasmic reticulum Ca2+-ATPase of fast twitch muscle from adult rabbit. It is generally acknowledged that the structure of SERCA1a is representative for the superfamily of P-type ATPases.
Hub AI
P-type ATPase AI simulator
(@P-type ATPase_simulator)
P-type ATPase
The P-type ATPases, also known as E1-E2 ATPases, are a large group of evolutionarily related ion and lipid pumps that are found in bacteria, archaea, and eukaryotes. P-type ATPases are α-helical bundle primary transporters named based upon their ability to catalyze auto- (or self-) phosphorylation (hence P) of a key conserved aspartate residue within the pump and their energy source, adenosine triphosphate (ATP). In addition, they all appear to interconvert between at least two different conformations, denoted by E1 and E2. P-type ATPases fall under the P-type ATPase (P-ATPase) Superfamily (TC# 3.A.3) which, as of early 2016, includes 20 different protein families.
Most members of this transporter superfamily drive cation uptake or efflux, though one subfamily, the flippases, is involved in flipping phospholipids to maintain the asymmetric nature of the biomembrane.
In humans, P-type ATPases serve as a basis for nerve impulses, relaxation of muscles, secretion and absorption in the kidney, absorption of nutrient in the intestine and other physiological processes. Prominent examples of P-type ATPases are the sodium-potassium pump (Na+/K+-ATPase), the proton-potassium pump (H+/K+-ATPase), the calcium pump (Ca2+-ATPase) and the plasma membrane proton pump (H+-ATPase) of plants and fungi.
The generalized reaction for P-type ATPases is
nLigand1 (out) + mLigand2 (in) + ATP → nLigand1 (in) + mLigand2 (out) + ADP + Pi.
where the ligand can be either a metal ion or a phospholipid molecule.
The first P-type ATPase discovered was the Na+/K+-ATPase, which Nobel laureate Jens Christian Skou isolated in 1957. The Na+/K+-ATPase was only the first member of a large and still-growing protein family (see Swiss-Prot Prosite motif PS00154).
P-type ATPases have a single catalytic subunit of 70 - 140 kDa. The catalytic subunit hydrolyzes ATP, contains the aspartyl phosphorylation site and binding sites for the transported ligand(s) and catalyzes ion transport. Various subfamilies of P-type ATPases also need additional subunits for proper function. Additional subunits that lack catalytic activity are present in the ATPase complexes of P1A, P2A, P2C and P4 ATPases. E.g. the catalytic alpha subunit of Na+/K+-ATPase consists of two additional subunits, beta and gamma, involved in trafficking, folding, and regulation of these pumps. The first P-type ATPase to be crystallized was SERCA1a, a sarco(endo)plasmic reticulum Ca2+-ATPase of fast twitch muscle from adult rabbit. It is generally acknowledged that the structure of SERCA1a is representative for the superfamily of P-type ATPases.