Community hub
Recent from talks
Contribute something to knowledge base
Content stats: 0 posts, 0 articles, 1 media, 0 notes
Members stats: 0 subscribers, 0 contributors, 0 moderators, 0 supporters
Subscribers
Supporters
Contributors
Moderators
Hub AI
Poliovirus AI simulator
(@Poliovirus_simulator)
Hub AI
Poliovirus AI simulator
(@Poliovirus_simulator)
Poliovirus
Poliovirus, the causative agent of polio (also known as poliomyelitis), is a serotype of the species Enterovirus C, in the family of Picornaviridae. There are three poliovirus serotypes, numbered 1, 2, and 3.
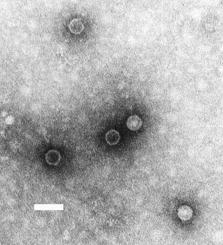
Poliovirus is composed of an RNA genome and a protein capsid. The genome is a single-stranded positive-sense RNA (+ssRNA) genome that is about 7500 nucleotides long. The viral particle is about 30 nm in diameter with icosahedral symmetry. Because of its short genome and its simple composition—only a strand of RNA and a nonenveloped icosahedral protein coat encapsulating it—poliovirus is widely regarded as the simplest significant virus.
Poliovirus is one of the most well-characterized viruses, and has become a useful model system for understanding the biology of RNA viruses.
Poliovirus infects human cells by binding to an immunoglobulin-like receptor, CD155 (also known as the poliovirus receptor or PVR) on the cell surface. Interaction of poliovirus and CD155 facilitates an irreversible conformational change of the viral particle necessary for viral entry. Following attachment to the host cell membrane, entry of the viral nucleic acid was thought to occur one of two ways: via the formation of a pore in the plasma membrane through which the RNA is then "injected" into the host cell cytoplasm, or via virus uptake by receptor-mediated endocytosis. Recent experimental evidence supports the latter hypothesis and suggests that poliovirus binds to CD155 and is taken up by endocytosis. Immediately after internalization of the particle, the viral RNA is released.
Poliovirus is a positive-stranded RNA virus. Thus, the genome enclosed within the viral particle can be used as messenger RNA and immediately translated by the host cell. On entry, the virus hijacks the cell's translation machinery, causing inhibition of cellular protein synthesis in favor of virus-specific protein production. Unlike the host cell's mRNAs, the 5' end of poliovirus RNA is extremely long—over 700 nucleotides—and highly structured. This region of the viral genome is called an internal ribosome entry site (IRES). This region consists of many secondary structures and 3 or 4 domains. Domain 3 is a self folding RNA element that contains conserved structural motifs in various stable stem loops linked by two four-way junctions. As IRES consists of many domains, these domains themselves consist of many loops that contribute to modified translation without a 5’ end cap by hijacking ribosomes. The interaction loop of domain 3 is known as GNRA tetraloop. The residues of adenosines A180 and A181 in the GUAA tetraloop form hydrogen bonds via non canonical base pairing interactions with the base pairs of the receptors C230/G242 and G231/C241, respectively. Genetic mutations in this region prevent viral protein production. The first IRES to be discovered was found in poliovirus RNA.
Poliovirus mRNA is translated as one long polypeptide. This polypeptide is then autocleaved by internal proteases into about 10 individual viral proteins. Not all cleavages occur with the same efficiency. Therefore, the amounts of proteins produced by the polypeptide cleavage vary: for example, smaller amounts of 3Dpol are produced than those of capsid proteins, VP1–4. These individual viral proteins are:
After translation, transcription and genome replication which involve a single process, synthesis of (+) RNA) is realized. For the infecting (+)RNA to be replicated, multiple copies of (−)RNA must be transcribed and then used as templates for (+)RNA synthesis. Replicative intermediates (RIs), which are an association of RNA molecules consisting of a template RNA and several growing RNAs of varying length, are seen in both the replication complexes for (−)RNAs and (+)RNAs. For synthesis of each negative-strand and positive-strand RNAs, VPg protein in the poliovirus works as a primer. RNA-dependent RNA polymerase of the poliovirus adds two uracil nucleotides (UU) to VPg protein utilizing the poly(A) tail at the 3′-end of the +ssRNA genome as a pattern for synthesis of the negative-strand antigenomic RNA. To initiate this −ssRNA synthesis, the tyrosine hydroxyl of VPg is needed. But for the initiation of positive strand RNA synthesis, CRE-dependent VPg uridylylation is needed. Which means that VPg is once more utilized as a primer however this time it adds the two uridine triphosphates using a cis-acting replication element (CRE) as a template.
The CRE of poliovirus is identified as an unachieved base-paired stem and a final loop consisting of 61 nt. The CRE is found in enteroviruses. It is a highly preserved secondary RNA structural element and bedded in the genome's polyprotein-coding region. The complex can be translocated to the 5' region of the genome that have no coding activity, at least 3.7-kb distant from the initial location. This process can occurs without negatively influencing activity. CRE copies do not influence replication negatively. Uridylylation process of VPg that takes place at CRE needs the presence of 3CDpro that is an RNA binding protein. It is attached to the CRE directly and specifically. Because of its presence VPg can bind the CRE properly and primary production proceeds without problems.
Poliovirus
Poliovirus, the causative agent of polio (also known as poliomyelitis), is a serotype of the species Enterovirus C, in the family of Picornaviridae. There are three poliovirus serotypes, numbered 1, 2, and 3.
Poliovirus is composed of an RNA genome and a protein capsid. The genome is a single-stranded positive-sense RNA (+ssRNA) genome that is about 7500 nucleotides long. The viral particle is about 30 nm in diameter with icosahedral symmetry. Because of its short genome and its simple composition—only a strand of RNA and a nonenveloped icosahedral protein coat encapsulating it—poliovirus is widely regarded as the simplest significant virus.
Poliovirus is one of the most well-characterized viruses, and has become a useful model system for understanding the biology of RNA viruses.
Poliovirus infects human cells by binding to an immunoglobulin-like receptor, CD155 (also known as the poliovirus receptor or PVR) on the cell surface. Interaction of poliovirus and CD155 facilitates an irreversible conformational change of the viral particle necessary for viral entry. Following attachment to the host cell membrane, entry of the viral nucleic acid was thought to occur one of two ways: via the formation of a pore in the plasma membrane through which the RNA is then "injected" into the host cell cytoplasm, or via virus uptake by receptor-mediated endocytosis. Recent experimental evidence supports the latter hypothesis and suggests that poliovirus binds to CD155 and is taken up by endocytosis. Immediately after internalization of the particle, the viral RNA is released.
Poliovirus is a positive-stranded RNA virus. Thus, the genome enclosed within the viral particle can be used as messenger RNA and immediately translated by the host cell. On entry, the virus hijacks the cell's translation machinery, causing inhibition of cellular protein synthesis in favor of virus-specific protein production. Unlike the host cell's mRNAs, the 5' end of poliovirus RNA is extremely long—over 700 nucleotides—and highly structured. This region of the viral genome is called an internal ribosome entry site (IRES). This region consists of many secondary structures and 3 or 4 domains. Domain 3 is a self folding RNA element that contains conserved structural motifs in various stable stem loops linked by two four-way junctions. As IRES consists of many domains, these domains themselves consist of many loops that contribute to modified translation without a 5’ end cap by hijacking ribosomes. The interaction loop of domain 3 is known as GNRA tetraloop. The residues of adenosines A180 and A181 in the GUAA tetraloop form hydrogen bonds via non canonical base pairing interactions with the base pairs of the receptors C230/G242 and G231/C241, respectively. Genetic mutations in this region prevent viral protein production. The first IRES to be discovered was found in poliovirus RNA.
Poliovirus mRNA is translated as one long polypeptide. This polypeptide is then autocleaved by internal proteases into about 10 individual viral proteins. Not all cleavages occur with the same efficiency. Therefore, the amounts of proteins produced by the polypeptide cleavage vary: for example, smaller amounts of 3Dpol are produced than those of capsid proteins, VP1–4. These individual viral proteins are:
After translation, transcription and genome replication which involve a single process, synthesis of (+) RNA) is realized. For the infecting (+)RNA to be replicated, multiple copies of (−)RNA must be transcribed and then used as templates for (+)RNA synthesis. Replicative intermediates (RIs), which are an association of RNA molecules consisting of a template RNA and several growing RNAs of varying length, are seen in both the replication complexes for (−)RNAs and (+)RNAs. For synthesis of each negative-strand and positive-strand RNAs, VPg protein in the poliovirus works as a primer. RNA-dependent RNA polymerase of the poliovirus adds two uracil nucleotides (UU) to VPg protein utilizing the poly(A) tail at the 3′-end of the +ssRNA genome as a pattern for synthesis of the negative-strand antigenomic RNA. To initiate this −ssRNA synthesis, the tyrosine hydroxyl of VPg is needed. But for the initiation of positive strand RNA synthesis, CRE-dependent VPg uridylylation is needed. Which means that VPg is once more utilized as a primer however this time it adds the two uridine triphosphates using a cis-acting replication element (CRE) as a template.
The CRE of poliovirus is identified as an unachieved base-paired stem and a final loop consisting of 61 nt. The CRE is found in enteroviruses. It is a highly preserved secondary RNA structural element and bedded in the genome's polyprotein-coding region. The complex can be translocated to the 5' region of the genome that have no coding activity, at least 3.7-kb distant from the initial location. This process can occurs without negatively influencing activity. CRE copies do not influence replication negatively. Uridylylation process of VPg that takes place at CRE needs the presence of 3CDpro that is an RNA binding protein. It is attached to the CRE directly and specifically. Because of its presence VPg can bind the CRE properly and primary production proceeds without problems.
Recent media
Recent media