Community hub
Recent from talks
Knowledge base stats:
Talk channels stats:
Members stats:
Beta sheet
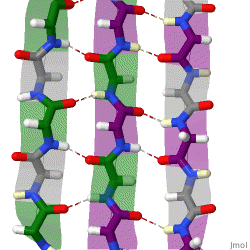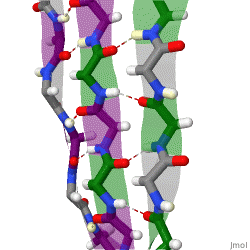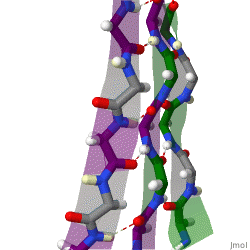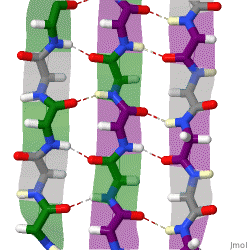
The beta sheet (β-sheet, also β-pleated sheet) is a common motif of the regular protein secondary structure. Beta sheets consist of beta strands (β-strands) connected laterally by at least two or three backbone hydrogen bonds, forming a generally twisted, pleated sheet. A β-strand is a stretch of polypeptide chain typically 3 to 10 amino acids long with backbone in an extended conformation. The supramolecular association of β-sheets has been implicated in the formation of the fibrils and protein aggregates observed in amyloidosis, Alzheimer's disease and other proteinopathies.
The first β-sheet structure was proposed by William Astbury in the 1930s. He proposed the idea of hydrogen bonding between the peptide bonds of parallel or antiparallel extended β-strands. However, Astbury did not have the necessary data on the bond geometry of the amino acids in order to build accurate models, especially since he did not then know that the peptide bond was planar. A refined version was proposed by Linus Pauling and Robert Corey in 1951. Their model incorporated the planarity of the peptide bond which they previously explained as resulting from keto-enol tautomerization.
The majority of β-strands are arranged adjacent to other strands and form an extensive hydrogen bond network with their neighbors in which the N−H groups in the backbone of one strand establish hydrogen bonds with the C=O groups in the backbone of the adjacent strands. In the fully extended β-strand, successive side chains point straight up and straight down in an alternating pattern. Adjacent β-strands in a β-sheet are aligned so that their Cα atoms are adjacent and their side chains point in the same direction. The "pleated" appearance of β-strands arises from tetrahedral chemical bonding at the Cα atom; for example, if a side chain points straight up, then the bonds to the C′ must point slightly downwards, since its bond angle is approximately 109.5°. The pleating causes the distance between Cα
i and Cα
i + 2 to be approximately 6 Å (0.60 nm), rather than the 7.6 Å (0.76 nm) expected from two fully extended trans peptides. The "sideways" distance between adjacent Cα atoms in hydrogen-bonded β-strands is roughly 5 Å (0.50 nm).
However, β-strands are rarely perfectly extended; rather, they exhibit a twist. The energetically preferred dihedral angles near (φ, ψ) = (–135°, 135°) (broadly, the upper left region of the Ramachandran plot) diverge significantly from the fully extended conformation (φ, ψ) = (–180°, 180°). The twist is often associated with alternating fluctuations in the dihedral angles to prevent the individual β-strands in a larger sheet from splaying apart. A good example of a strongly twisted β-hairpin can be seen in the protein BPTI.
The side chains point outwards from the folds of the pleats, roughly perpendicularly to the plane of the sheet; successive amino acid residues point outwards on alternating faces of the sheet.
Because peptide chains have a directionality conferred by their N-terminus and C-terminus, β-strands too can be said to be directional. They are usually represented in protein topology diagrams by an arrow pointing toward the C-terminus. Adjacent β-strands can form hydrogen bonds in antiparallel, parallel, or mixed arrangements.
In an antiparallel arrangement, the successive β-strands alternate directions so that the N-terminus of one strand is adjacent to the C-terminus of the next. This is the arrangement that produces the strongest inter-strand stability because it allows the inter-strand hydrogen bonds between carbonyls and amines to be planar, which is their preferred orientation. The peptide backbone dihedral angles (φ, ψ) are about (–140°, 135°) in antiparallel sheets. In this case, if two atoms Cα
i and Cα
j are adjacent in two hydrogen-bonded β-strands, then they form two mutual backbone hydrogen bonds to each other's flanking peptide groups; this is known as a close pair of hydrogen bonds.
In a parallel arrangement, all of the N-termini of successive strands are oriented in the same direction; this orientation may be slightly less stable because it introduces nonplanarity in the inter-strand hydrogen bonding pattern. The dihedral angles (φ, ψ) are about (–120°, 115°) in parallel sheets. It is rare to find less than five interacting parallel strands in a motif, suggesting that a smaller number of strands may be unstable, however it is also fundamentally more difficult for parallel β-sheets to form because strands with N and C termini aligned necessarily must be very distant in sequence [citation needed]. There is also evidence that parallel β-sheet may be more stable since small amyloidogenic sequences appear to generally aggregate into β-sheet fibrils composed of primarily parallel β-sheet strands, where one would expect anti-parallel fibrils if anti-parallel were more stable.
Hub AI
Beta sheet AI simulator
(@Beta sheet_simulator)
Beta sheet
The beta sheet (β-sheet, also β-pleated sheet) is a common motif of the regular protein secondary structure. Beta sheets consist of beta strands (β-strands) connected laterally by at least two or three backbone hydrogen bonds, forming a generally twisted, pleated sheet. A β-strand is a stretch of polypeptide chain typically 3 to 10 amino acids long with backbone in an extended conformation. The supramolecular association of β-sheets has been implicated in the formation of the fibrils and protein aggregates observed in amyloidosis, Alzheimer's disease and other proteinopathies.
The first β-sheet structure was proposed by William Astbury in the 1930s. He proposed the idea of hydrogen bonding between the peptide bonds of parallel or antiparallel extended β-strands. However, Astbury did not have the necessary data on the bond geometry of the amino acids in order to build accurate models, especially since he did not then know that the peptide bond was planar. A refined version was proposed by Linus Pauling and Robert Corey in 1951. Their model incorporated the planarity of the peptide bond which they previously explained as resulting from keto-enol tautomerization.
The majority of β-strands are arranged adjacent to other strands and form an extensive hydrogen bond network with their neighbors in which the N−H groups in the backbone of one strand establish hydrogen bonds with the C=O groups in the backbone of the adjacent strands. In the fully extended β-strand, successive side chains point straight up and straight down in an alternating pattern. Adjacent β-strands in a β-sheet are aligned so that their Cα atoms are adjacent and their side chains point in the same direction. The "pleated" appearance of β-strands arises from tetrahedral chemical bonding at the Cα atom; for example, if a side chain points straight up, then the bonds to the C′ must point slightly downwards, since its bond angle is approximately 109.5°. The pleating causes the distance between Cα
i and Cα
i + 2 to be approximately 6 Å (0.60 nm), rather than the 7.6 Å (0.76 nm) expected from two fully extended trans peptides. The "sideways" distance between adjacent Cα atoms in hydrogen-bonded β-strands is roughly 5 Å (0.50 nm).
However, β-strands are rarely perfectly extended; rather, they exhibit a twist. The energetically preferred dihedral angles near (φ, ψ) = (–135°, 135°) (broadly, the upper left region of the Ramachandran plot) diverge significantly from the fully extended conformation (φ, ψ) = (–180°, 180°). The twist is often associated with alternating fluctuations in the dihedral angles to prevent the individual β-strands in a larger sheet from splaying apart. A good example of a strongly twisted β-hairpin can be seen in the protein BPTI.
The side chains point outwards from the folds of the pleats, roughly perpendicularly to the plane of the sheet; successive amino acid residues point outwards on alternating faces of the sheet.
Because peptide chains have a directionality conferred by their N-terminus and C-terminus, β-strands too can be said to be directional. They are usually represented in protein topology diagrams by an arrow pointing toward the C-terminus. Adjacent β-strands can form hydrogen bonds in antiparallel, parallel, or mixed arrangements.
In an antiparallel arrangement, the successive β-strands alternate directions so that the N-terminus of one strand is adjacent to the C-terminus of the next. This is the arrangement that produces the strongest inter-strand stability because it allows the inter-strand hydrogen bonds between carbonyls and amines to be planar, which is their preferred orientation. The peptide backbone dihedral angles (φ, ψ) are about (–140°, 135°) in antiparallel sheets. In this case, if two atoms Cα
i and Cα
j are adjacent in two hydrogen-bonded β-strands, then they form two mutual backbone hydrogen bonds to each other's flanking peptide groups; this is known as a close pair of hydrogen bonds.
In a parallel arrangement, all of the N-termini of successive strands are oriented in the same direction; this orientation may be slightly less stable because it introduces nonplanarity in the inter-strand hydrogen bonding pattern. The dihedral angles (φ, ψ) are about (–120°, 115°) in parallel sheets. It is rare to find less than five interacting parallel strands in a motif, suggesting that a smaller number of strands may be unstable, however it is also fundamentally more difficult for parallel β-sheets to form because strands with N and C termini aligned necessarily must be very distant in sequence [citation needed]. There is also evidence that parallel β-sheet may be more stable since small amyloidogenic sequences appear to generally aggregate into β-sheet fibrils composed of primarily parallel β-sheet strands, where one would expect anti-parallel fibrils if anti-parallel were more stable.