
Community hub
Recent from talks
Contribute something to knowledge base
Content stats: 0 posts, 0 articles, 1 media, 0 notes
Members stats: 0 subscribers, 0 contributors, 0 moderators, 0 supporters
Subscribers
Supporters
Contributors
Moderators
Hub AI
Entorhinal cortex AI simulator
(@Entorhinal cortex_simulator)
Hub AI
Entorhinal cortex AI simulator
(@Entorhinal cortex_simulator)
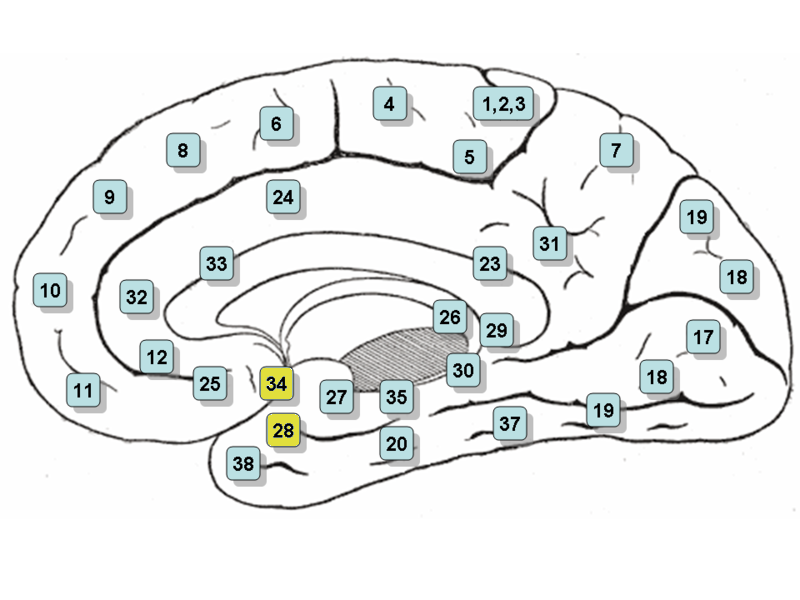
Entorhinal cortex
The entorhinal cortex (EC) is an area of the brain's allocortex, located in the medial temporal lobe, whose functions include being a widespread network hub for memory, navigation, and the perception of time. The EC is the main interface between the hippocampus and neocortex. The EC-hippocampus system plays an important role in declarative (autobiographical/episodic/semantic) memories and in particular spatial memories including memory formation, memory consolidation, and memory optimization in sleep. The EC is also responsible for the pre-processing (familiarity) of the input signals in the reflex nictitating membrane response of classical trace conditioning; the association of impulses from the eye and the ear occurs in the entorhinal cortex.
The entorhinal cortex is a portion of the rostral parahippocampal gyrus.
It is usually divided into medial and lateral regions with three bands with distinct properties and connectivity running perpendicular across the whole area. A distinguishing characteristic of the EC is the lack of cell bodies where layer IV should be; this layer is called the Lamina dissecans.
The superficial layers – layers II and III – of EC project to the dentate gyrus and hippocampus: Layer II projects primarily to dentate gyrus and hippocampal region CA3; layer III projects primarily to hippocampal region CA1 and the subiculum. These layers receive input from other cortical areas, especially associational, perirhinal, and parahippocampal cortices, as well as prefrontal cortex. EC as a whole, therefore, receives highly processed input from every sensory modality, as well as input relating to ongoing cognitive processes, though it should be stressed that, within EC, this information remains at least partially segregated.
The deep layers, especially layer V, receive one of the three main outputs of the hippocampus and, in turn, reciprocate connections from other cortical areas that project to superficial EC.
In 2005, it was discovered that entorhinal cortex contains a neural map of the spatial environment in rats. In 2014, John O'Keefe, May-Britt Moser and Edvard Moser received the Nobel Prize in Physiology or Medicine, partly because of this discovery.
In rodents, neurons in the lateral entorhinal cortex exhibit little spatial selectivity, whereas neurons of the medial entorhinal cortex (MEC), exhibit multiple "place fields" that are arranged in a hexagonal pattern, and are, therefore, called "grid cells". These fields and spacing between fields increase from the dorso-lateral MEA to the ventro-medial MEA.
The same group of researchers has found speed cells in the medial entorhinal cortex of rats. The speed of movement is translated from proprioceptive information and is represented as firing rates in these cells. The cells are known to fire in correlation to future speed of the rodent.
Entorhinal cortex
The entorhinal cortex (EC) is an area of the brain's allocortex, located in the medial temporal lobe, whose functions include being a widespread network hub for memory, navigation, and the perception of time. The EC is the main interface between the hippocampus and neocortex. The EC-hippocampus system plays an important role in declarative (autobiographical/episodic/semantic) memories and in particular spatial memories including memory formation, memory consolidation, and memory optimization in sleep. The EC is also responsible for the pre-processing (familiarity) of the input signals in the reflex nictitating membrane response of classical trace conditioning; the association of impulses from the eye and the ear occurs in the entorhinal cortex.
The entorhinal cortex is a portion of the rostral parahippocampal gyrus.
It is usually divided into medial and lateral regions with three bands with distinct properties and connectivity running perpendicular across the whole area. A distinguishing characteristic of the EC is the lack of cell bodies where layer IV should be; this layer is called the Lamina dissecans.
The superficial layers – layers II and III – of EC project to the dentate gyrus and hippocampus: Layer II projects primarily to dentate gyrus and hippocampal region CA3; layer III projects primarily to hippocampal region CA1 and the subiculum. These layers receive input from other cortical areas, especially associational, perirhinal, and parahippocampal cortices, as well as prefrontal cortex. EC as a whole, therefore, receives highly processed input from every sensory modality, as well as input relating to ongoing cognitive processes, though it should be stressed that, within EC, this information remains at least partially segregated.
The deep layers, especially layer V, receive one of the three main outputs of the hippocampus and, in turn, reciprocate connections from other cortical areas that project to superficial EC.
In 2005, it was discovered that entorhinal cortex contains a neural map of the spatial environment in rats. In 2014, John O'Keefe, May-Britt Moser and Edvard Moser received the Nobel Prize in Physiology or Medicine, partly because of this discovery.
In rodents, neurons in the lateral entorhinal cortex exhibit little spatial selectivity, whereas neurons of the medial entorhinal cortex (MEC), exhibit multiple "place fields" that are arranged in a hexagonal pattern, and are, therefore, called "grid cells". These fields and spacing between fields increase from the dorso-lateral MEA to the ventro-medial MEA.
The same group of researchers has found speed cells in the medial entorhinal cortex of rats. The speed of movement is translated from proprioceptive information and is represented as firing rates in these cells. The cells are known to fire in correlation to future speed of the rodent.
Recent media
Recent media