Recent from talks
Neocortex
Knowledge base stats:
Talk channels stats:
Members stats:
Neocortex
The neocortex, also called the neopallium, isocortex, or the six-layered cortex, is a set of layers of the mammalian cerebral cortex involved in higher-order brain functions such as sensory perception, cognition, generation of motor commands, spatial reasoning, and language. The neocortex is further subdivided into the true isocortex and the proisocortex.
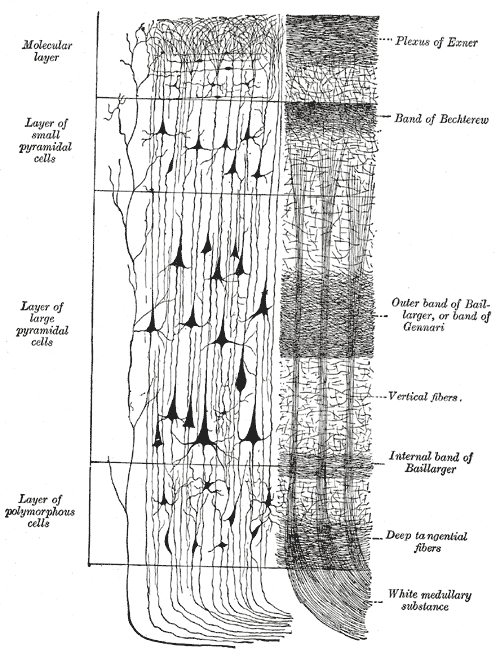
In the human brain, the cerebral cortex consists of the larger neocortex and the smaller allocortex, respectively taking up 90% and 10%. The neocortex is made up of six layers, labelled from the outermost inwards, I to VI.
The term is from cortex, Latin, "bark" or "rind", combined with neo-, Greek, "new". Neopallium is a similar hybrid, from Latin pallium, "cloak". Isocortex and allocortex are hybrids with Greek isos, "same", and allos, "other".
The neocortex is the most developed in its organisation and number of layers, of the cerebral tissues. The neocortex consists of the grey matter, or neuronal cell bodies and unmyelinated fibers, surrounding the deeper white matter (myelinated axons) in the cerebrum. This is a very thin layer though, about 2–4 mm thick. There are two types of cortex in the neocortex, the proisocortex and the true isocortex. The pro-isocortex is a transitional area between the true isocortex and the periallocortex (part of the allocortex). It is found in the cingulate cortex (part of the limbic system), in Brodmann's areas 24, 25, 30 and 32, the insula and the parahippocampal gyrus.
Of all the mammals studied to date (including humans), a species of oceanic dolphin known as the long-finned pilot whale has been found to have the most neocortical neurons.
The neocortex is smooth in rodents and other small mammals, whereas in elephants, dolphins and primates and other larger mammals it has deep grooves (sulci) and ridges (gyri). These folds allow the surface area of the neocortex to be greatly increased. All human brains have the same overall pattern of main gyri and sulci, although they differ in detail from one person to another. The mechanism by which the gyri form during embryogenesis is not entirely clear, and there are several competing hypotheses that explain gyrification, such as axonal tension, cortical buckling or differences in cellular proliferation rates in different areas of the cortex.
The neocortex contains both excitatory (~80%) and inhibitory (~20%) neurons, named for their effect on other neurons. The human neocortex consists of hundreds of different types of cells. The structure of the neocortex is relatively uniform (hence the alternative names "iso-" and "homotypic" cortex), consisting of six horizontal layers segregated principally by cell type and neuronal connections. However, there are many exceptions to this uniformity; for example, layer IV is small or missing in the primary motor cortex. There is some canonical circuitry within the cortex; for example, pyramidal neurons in the upper layers II and III project their axons to other areas of neocortex, while those in the deeper layers V and VI often project out of the cortex, e.g. to the thalamus, brainstem, and spinal cord. Neurons in layer IV receive the majority of the synaptic connections from outside the cortex (mostly from thalamus), and themselves make short-range, local connections to other cortical layers. Thus, layer IV is the main recipient of incoming sensory information and distributes it to the other layers for further processing.
The neocortex is often described as being arranged in vertical structures called cortical columns, patches of neocortex with a diameter of roughly 0.5 mm (and a depth of 2 mm, i.e., spanning all six layers). These columns are often thought of as the basic repeating functional units of the neocortex, but their many definitions, in terms of anatomy, size, or function, are generally not consistent with each other, leading to a lack of consensus regarding their structure or function or even whether it makes sense to try to understand the neocortex in terms of columns.
Hub AI
Neocortex AI simulator
(@Neocortex_simulator)
Neocortex
The neocortex, also called the neopallium, isocortex, or the six-layered cortex, is a set of layers of the mammalian cerebral cortex involved in higher-order brain functions such as sensory perception, cognition, generation of motor commands, spatial reasoning, and language. The neocortex is further subdivided into the true isocortex and the proisocortex.
In the human brain, the cerebral cortex consists of the larger neocortex and the smaller allocortex, respectively taking up 90% and 10%. The neocortex is made up of six layers, labelled from the outermost inwards, I to VI.
The term is from cortex, Latin, "bark" or "rind", combined with neo-, Greek, "new". Neopallium is a similar hybrid, from Latin pallium, "cloak". Isocortex and allocortex are hybrids with Greek isos, "same", and allos, "other".
The neocortex is the most developed in its organisation and number of layers, of the cerebral tissues. The neocortex consists of the grey matter, or neuronal cell bodies and unmyelinated fibers, surrounding the deeper white matter (myelinated axons) in the cerebrum. This is a very thin layer though, about 2–4 mm thick. There are two types of cortex in the neocortex, the proisocortex and the true isocortex. The pro-isocortex is a transitional area between the true isocortex and the periallocortex (part of the allocortex). It is found in the cingulate cortex (part of the limbic system), in Brodmann's areas 24, 25, 30 and 32, the insula and the parahippocampal gyrus.
Of all the mammals studied to date (including humans), a species of oceanic dolphin known as the long-finned pilot whale has been found to have the most neocortical neurons.
The neocortex is smooth in rodents and other small mammals, whereas in elephants, dolphins and primates and other larger mammals it has deep grooves (sulci) and ridges (gyri). These folds allow the surface area of the neocortex to be greatly increased. All human brains have the same overall pattern of main gyri and sulci, although they differ in detail from one person to another. The mechanism by which the gyri form during embryogenesis is not entirely clear, and there are several competing hypotheses that explain gyrification, such as axonal tension, cortical buckling or differences in cellular proliferation rates in different areas of the cortex.
The neocortex contains both excitatory (~80%) and inhibitory (~20%) neurons, named for their effect on other neurons. The human neocortex consists of hundreds of different types of cells. The structure of the neocortex is relatively uniform (hence the alternative names "iso-" and "homotypic" cortex), consisting of six horizontal layers segregated principally by cell type and neuronal connections. However, there are many exceptions to this uniformity; for example, layer IV is small or missing in the primary motor cortex. There is some canonical circuitry within the cortex; for example, pyramidal neurons in the upper layers II and III project their axons to other areas of neocortex, while those in the deeper layers V and VI often project out of the cortex, e.g. to the thalamus, brainstem, and spinal cord. Neurons in layer IV receive the majority of the synaptic connections from outside the cortex (mostly from thalamus), and themselves make short-range, local connections to other cortical layers. Thus, layer IV is the main recipient of incoming sensory information and distributes it to the other layers for further processing.
The neocortex is often described as being arranged in vertical structures called cortical columns, patches of neocortex with a diameter of roughly 0.5 mm (and a depth of 2 mm, i.e., spanning all six layers). These columns are often thought of as the basic repeating functional units of the neocortex, but their many definitions, in terms of anatomy, size, or function, are generally not consistent with each other, leading to a lack of consensus regarding their structure or function or even whether it makes sense to try to understand the neocortex in terms of columns.
Recent media