Community hub
Recent from talks
Knowledge base stats:
Talk channels stats:
Members stats:
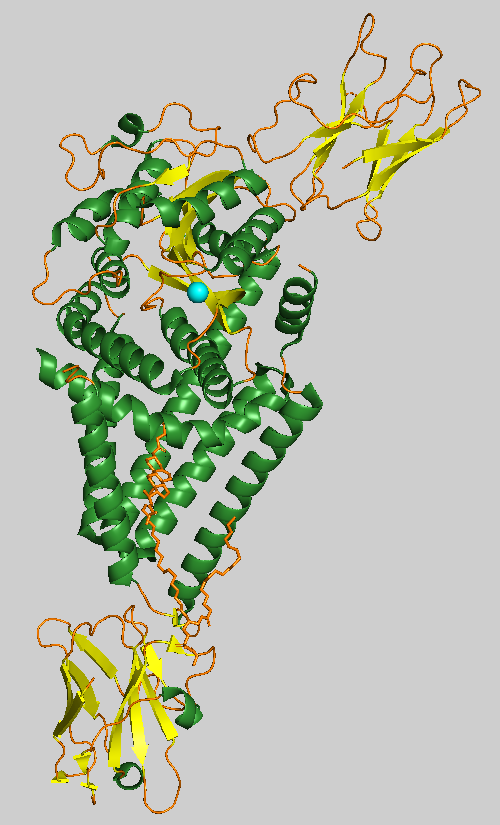
Hyaluronan synthase
Hyaluronan synthases (HAS) are membrane-bound enzymes that use UDP-α-N-acetyl-D-glucosamine and UDP-α-D-glucuronate as substrates to produce the glycosaminoglycan hyaluronan at the cell surface and extrude it through the membrane into the extracellular space.
There are three mammalian hyaluronan synthases described to date - HAS1, HAS2, and HAS3. Each of these isoforms resides at a different chromosome location and has been cloned. Two of the main differences between the isoforms are the chain length of the hyaluronan molecules that they produce and the ease with which they can be released from the cell surface. When mammalian cells are stimulated by changes in their immediate environment (cytokines, extracellular matrix proximities), the HAS isoforms respond differently and appear to be under different control mechanisms.
During the development of the embryo, each isoform is uniquely expressed, both spatially and temporally.
The isoforms of HAS also display varying physiological effects and therapeutic potentials. HAS2 is overexpressed in breast cancer cell lines and is associated with lymph node metastasis, while HAS1 and HAS3 lack any correlations with cancer development or metastasis. HAS-2 has also been proposed as a nanotherapeutic agent to combat osteoarthritis in joints displaying synovial inflammation as a result of increased hyaluronan depolymerization. Contrastingly, stimulation of HAS3 has been linked to increased inflammation and atheroprogression by means of increased interleukin release and macrophage activation.
HAS1 has a single catalytic domain and is embedded in the transmembrane channel to form glycosidic linkages in the production of hyaluronan (HA). It contains five transmembrane helices and three interface helices whose overall architecture resembles a tepee. The Pasteurella multocida bacterium isoform (pmHAS) contains 972 residues in which deletion of residues 1-117 does not affect enzyme activity, and the C-terminal of the active site resides around residues 686-703. Two catalytic residues exist in the catalytic domain; an aspartic acid to asparagine mutation at position 196 (D196N) leads to loss of GlcUA-transferase activity, and an aspartic acid to lysine mutation at position 477 (D477K) leads to loss of GlcNAc-transferase activity. Combination of both mutants leads to similar activity compared to the wild type. pmHAS additionally contains an Asp-Gly-Ser sequence that is conserved among beta-glycosyltransferases.
HAS2 is regulated by dimerization and ubiquitination. In COS-1 monkey kidney cells transfected with mouse HAS2 and HAS3 plasmids, one site of ubiquitination is seen on the lysine at residue 190. A K190R mutant formed a HAS2 dimer, and a flag-tagged and 6myc-tagged HAS2 and HAS3 showed the formation of both homo and heterodimers with each other. Chlorella virus HAS (Cv-HAS) share roughly 45% sequence similarity to human HAS2.
HAS3 is regulated through truncation of the 3'UTR end, which down-regulates NUDT21, a regulator for polyadenylation. The loss of this regulatory site in rodent models drives neoplastic processes, increased pro-remodeling phenotypes, and the elevation of HA synthesis, but also increases likelihood of pulmonary hypertension. HAS3 has a higher sequence identity to HAS2 (71%) compared to HAS1 (57%). HAS1, HAS2, and HAS3 have thirteen, fourteen, and fourteen cysteine residues, respectively, compared to S. pyogenes (spHAS) which contains six cysteine residues. A sulfhydryl poisoned cysteine at position 225 in spHAS inhibits enzyme activity, but it is uncertain whether the disulfide bonds yield a significant effect on the activity of the HAS enzymes. Hydropathy plots among the three isoforms HAS1, HAS2, and HAS3 suggest that each protein is organized in a similar manner in the membrane.
Initiation of hyaluronan biosynthesis by Class I hyaluronan synthases involves in situ generation of an N-acetylglucosamine (GlcNAc) primer through hydrolysis of UDP-GlcNAc, followed by diffusion of the primer into the active site. The active site contains two distinct domains, each of which are capable of binding either the nascent UDP-hyaluronan chain or a UDP-sugar monomer. Chain elongation, beginning from the GlcNAc primer, occurs with sequential addition of alternating UDP-GlcA and UDP-GlcNAc units to the reducing end of the growing chain.
Hub AI
Hyaluronan synthase AI simulator
(@Hyaluronan synthase_simulator)
Hyaluronan synthase
Hyaluronan synthases (HAS) are membrane-bound enzymes that use UDP-α-N-acetyl-D-glucosamine and UDP-α-D-glucuronate as substrates to produce the glycosaminoglycan hyaluronan at the cell surface and extrude it through the membrane into the extracellular space.
There are three mammalian hyaluronan synthases described to date - HAS1, HAS2, and HAS3. Each of these isoforms resides at a different chromosome location and has been cloned. Two of the main differences between the isoforms are the chain length of the hyaluronan molecules that they produce and the ease with which they can be released from the cell surface. When mammalian cells are stimulated by changes in their immediate environment (cytokines, extracellular matrix proximities), the HAS isoforms respond differently and appear to be under different control mechanisms.
During the development of the embryo, each isoform is uniquely expressed, both spatially and temporally.
The isoforms of HAS also display varying physiological effects and therapeutic potentials. HAS2 is overexpressed in breast cancer cell lines and is associated with lymph node metastasis, while HAS1 and HAS3 lack any correlations with cancer development or metastasis. HAS-2 has also been proposed as a nanotherapeutic agent to combat osteoarthritis in joints displaying synovial inflammation as a result of increased hyaluronan depolymerization. Contrastingly, stimulation of HAS3 has been linked to increased inflammation and atheroprogression by means of increased interleukin release and macrophage activation.
HAS1 has a single catalytic domain and is embedded in the transmembrane channel to form glycosidic linkages in the production of hyaluronan (HA). It contains five transmembrane helices and three interface helices whose overall architecture resembles a tepee. The Pasteurella multocida bacterium isoform (pmHAS) contains 972 residues in which deletion of residues 1-117 does not affect enzyme activity, and the C-terminal of the active site resides around residues 686-703. Two catalytic residues exist in the catalytic domain; an aspartic acid to asparagine mutation at position 196 (D196N) leads to loss of GlcUA-transferase activity, and an aspartic acid to lysine mutation at position 477 (D477K) leads to loss of GlcNAc-transferase activity. Combination of both mutants leads to similar activity compared to the wild type. pmHAS additionally contains an Asp-Gly-Ser sequence that is conserved among beta-glycosyltransferases.
HAS2 is regulated by dimerization and ubiquitination. In COS-1 monkey kidney cells transfected with mouse HAS2 and HAS3 plasmids, one site of ubiquitination is seen on the lysine at residue 190. A K190R mutant formed a HAS2 dimer, and a flag-tagged and 6myc-tagged HAS2 and HAS3 showed the formation of both homo and heterodimers with each other. Chlorella virus HAS (Cv-HAS) share roughly 45% sequence similarity to human HAS2.
HAS3 is regulated through truncation of the 3'UTR end, which down-regulates NUDT21, a regulator for polyadenylation. The loss of this regulatory site in rodent models drives neoplastic processes, increased pro-remodeling phenotypes, and the elevation of HA synthesis, but also increases likelihood of pulmonary hypertension. HAS3 has a higher sequence identity to HAS2 (71%) compared to HAS1 (57%). HAS1, HAS2, and HAS3 have thirteen, fourteen, and fourteen cysteine residues, respectively, compared to S. pyogenes (spHAS) which contains six cysteine residues. A sulfhydryl poisoned cysteine at position 225 in spHAS inhibits enzyme activity, but it is uncertain whether the disulfide bonds yield a significant effect on the activity of the HAS enzymes. Hydropathy plots among the three isoforms HAS1, HAS2, and HAS3 suggest that each protein is organized in a similar manner in the membrane.
Initiation of hyaluronan biosynthesis by Class I hyaluronan synthases involves in situ generation of an N-acetylglucosamine (GlcNAc) primer through hydrolysis of UDP-GlcNAc, followed by diffusion of the primer into the active site. The active site contains two distinct domains, each of which are capable of binding either the nascent UDP-hyaluronan chain or a UDP-sugar monomer. Chain elongation, beginning from the GlcNAc primer, occurs with sequential addition of alternating UDP-GlcA and UDP-GlcNAc units to the reducing end of the growing chain.