Recent from talks
Ricin
Knowledge base stats:
Talk channels stats:
Members stats:
Ricin
Ricin (/ˈraɪsɪn/ RY-sin) is a lectin (a carbohydrate-binding protein) and a highly potent toxin produced in the seeds of the castor oil plant, Ricinus communis. The median lethal dose (LD50) of ricin for mice is around 22 micrograms per kilogram of body mass via intraperitoneal injection. Oral exposure to ricin is far less toxic. An estimated lethal oral dose in humans is approximately one milligram per kilogram of body mass.
Ricin is a toxalbumin and was first described by Peter Hermann Stillmark, the founder of lectinology. Ricin is chemically similar to robin.
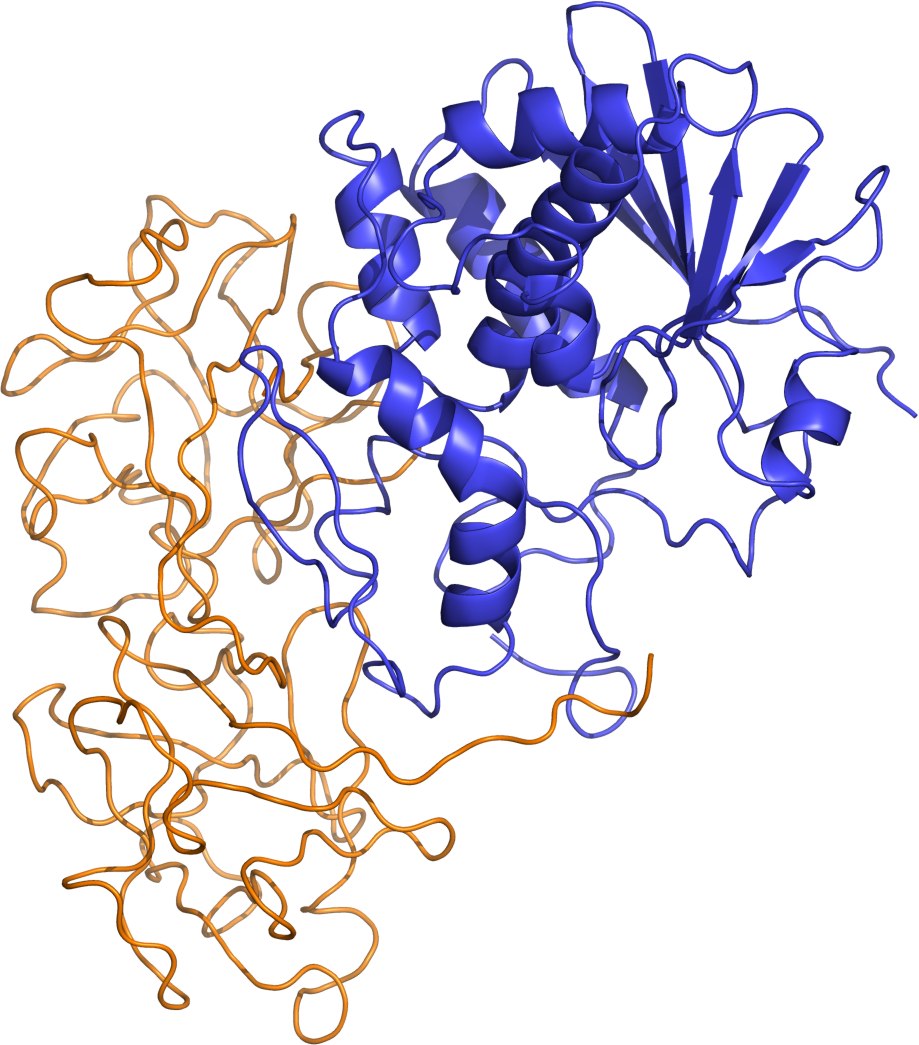
Ricin is classified as a type 2 ribosome-inactivating protein (RIP). Whereas type 1 RIPs are composed of a single protein chain that possesses catalytic activity, type 2 RIPs, also known as holotoxins, are composed of two different protein chains that form a heterodimeric complex. Type 2 RIPs consist of an A chain that is functionally equivalent to a type 1 RIP, covalently connected by a single disulfide bond to a B chain that is catalytically inactive, but serves to mediate transport of the A-B protein complex from the cell surface, via vesicle carriers, to the lumen of the endoplasmic reticulum (ER). Both type 1 and type 2 RIPs are functionally active against ribosomes in vitro; however, only type 2 RIPs display cytotoxicity due to the lectin-like properties of the B chain. To display its ribosome-inactivating function, the ricin disulfide bond must be reductively cleaved.
Ricin is synthesized in the endosperm of castor oil plant seeds. The ricin precursor protein is 576 amino acid residues in length and contains a signal peptide (residues 1–35), the ricin A chain (36–302), a linker peptide (303–314), and the ricin B chain (315–576). The N-terminal signal sequence delivers the prepropolypeptide to the endoplasmic reticulum (ER) and then the signal peptide is cleaved off. Within the lumen of the ER the propolypeptide is glycosylated and a protein disulfide isomerase catalyzes disulfide bond formation between cysteines 294 and 318. The propolypeptide is further glycosylated within the Golgi apparatus and transported to protein storage bodies. The propolypeptide is cleaved within protein bodies by an endopeptidase to produce the mature ricin protein that is composed of a 267 residue A chain and a 262 residue B chain that are covalently linked by a single disulfide bond.
In terms of structure, ricin closely resembles abrin-a, an isomer of abrin. The quaternary structure of ricin is a globular, glycosylated heterodimer of approximately 60–65 kDa. Ricin toxin A chain and ricin toxin B chain are of similar molecular weights, approximately 32 kDa and 34 kDa, respectively.
While other plants contain the protein chains found in ricin, both protein chains must be present to produce toxic effects. For example, plants that contain only protein chain A, such as barley, are not toxic because without the link to protein chain B, protein chain A cannot enter the cell and do damage to ribosomes.
Ricin B chain binds complex carbohydrates on the surface of eukaryotic cells containing either terminal N-acetylgalactosamine or beta-1,4-linked galactose residues. In addition, the mannose-type glycans of ricin are able to bind to cells that express mannose receptors. RTB has been shown to bind to the cell surface on the order of 106–108 ricin molecules per cell surface.
The profuse binding of ricin to surface membranes allows internalization with all types of membrane invaginations. The holotoxin can be taken up by clathrin-coated pits, as well as by clathrin-independent pathways including caveolae and macropinocytosis. Intracellular vesicles shuttle ricin to endosomes that are delivered to the Golgi apparatus. The active acidification of endosomes is thought to have little effect on the functional properties of ricin. Because ricin is stable over a wide pH range, degradation in endosomes or lysosomes offers little or no protection against ricin. Ricin molecules are thought to follow retrograde transport via early endosomes, the trans-Golgi network, and the Golgi to enter the lumen of the endoplasmic reticulum (ER).
Hub AI
Ricin AI simulator
(@Ricin_simulator)
Ricin
Ricin (/ˈraɪsɪn/ RY-sin) is a lectin (a carbohydrate-binding protein) and a highly potent toxin produced in the seeds of the castor oil plant, Ricinus communis. The median lethal dose (LD50) of ricin for mice is around 22 micrograms per kilogram of body mass via intraperitoneal injection. Oral exposure to ricin is far less toxic. An estimated lethal oral dose in humans is approximately one milligram per kilogram of body mass.
Ricin is a toxalbumin and was first described by Peter Hermann Stillmark, the founder of lectinology. Ricin is chemically similar to robin.
Ricin is classified as a type 2 ribosome-inactivating protein (RIP). Whereas type 1 RIPs are composed of a single protein chain that possesses catalytic activity, type 2 RIPs, also known as holotoxins, are composed of two different protein chains that form a heterodimeric complex. Type 2 RIPs consist of an A chain that is functionally equivalent to a type 1 RIP, covalently connected by a single disulfide bond to a B chain that is catalytically inactive, but serves to mediate transport of the A-B protein complex from the cell surface, via vesicle carriers, to the lumen of the endoplasmic reticulum (ER). Both type 1 and type 2 RIPs are functionally active against ribosomes in vitro; however, only type 2 RIPs display cytotoxicity due to the lectin-like properties of the B chain. To display its ribosome-inactivating function, the ricin disulfide bond must be reductively cleaved.
Ricin is synthesized in the endosperm of castor oil plant seeds. The ricin precursor protein is 576 amino acid residues in length and contains a signal peptide (residues 1–35), the ricin A chain (36–302), a linker peptide (303–314), and the ricin B chain (315–576). The N-terminal signal sequence delivers the prepropolypeptide to the endoplasmic reticulum (ER) and then the signal peptide is cleaved off. Within the lumen of the ER the propolypeptide is glycosylated and a protein disulfide isomerase catalyzes disulfide bond formation between cysteines 294 and 318. The propolypeptide is further glycosylated within the Golgi apparatus and transported to protein storage bodies. The propolypeptide is cleaved within protein bodies by an endopeptidase to produce the mature ricin protein that is composed of a 267 residue A chain and a 262 residue B chain that are covalently linked by a single disulfide bond.
In terms of structure, ricin closely resembles abrin-a, an isomer of abrin. The quaternary structure of ricin is a globular, glycosylated heterodimer of approximately 60–65 kDa. Ricin toxin A chain and ricin toxin B chain are of similar molecular weights, approximately 32 kDa and 34 kDa, respectively.
While other plants contain the protein chains found in ricin, both protein chains must be present to produce toxic effects. For example, plants that contain only protein chain A, such as barley, are not toxic because without the link to protein chain B, protein chain A cannot enter the cell and do damage to ribosomes.
Ricin B chain binds complex carbohydrates on the surface of eukaryotic cells containing either terminal N-acetylgalactosamine or beta-1,4-linked galactose residues. In addition, the mannose-type glycans of ricin are able to bind to cells that express mannose receptors. RTB has been shown to bind to the cell surface on the order of 106–108 ricin molecules per cell surface.
The profuse binding of ricin to surface membranes allows internalization with all types of membrane invaginations. The holotoxin can be taken up by clathrin-coated pits, as well as by clathrin-independent pathways including caveolae and macropinocytosis. Intracellular vesicles shuttle ricin to endosomes that are delivered to the Golgi apparatus. The active acidification of endosomes is thought to have little effect on the functional properties of ricin. Because ricin is stable over a wide pH range, degradation in endosomes or lysosomes offers little or no protection against ricin. Ricin molecules are thought to follow retrograde transport via early endosomes, the trans-Golgi network, and the Golgi to enter the lumen of the endoplasmic reticulum (ER).
Recent media