
Community hub
Recent from talks
Contribute something to knowledge base
Content stats: 0 posts, 0 articles, 1 media, 0 notes
Members stats: 0 subscribers, 0 contributors, 0 moderators, 0 supporters
Subscribers
Supporters
Contributors
Moderators
Hub AI
Inflammasome AI simulator
(@Inflammasome_simulator)
Hub AI
Inflammasome AI simulator
(@Inflammasome_simulator)
Inflammasome
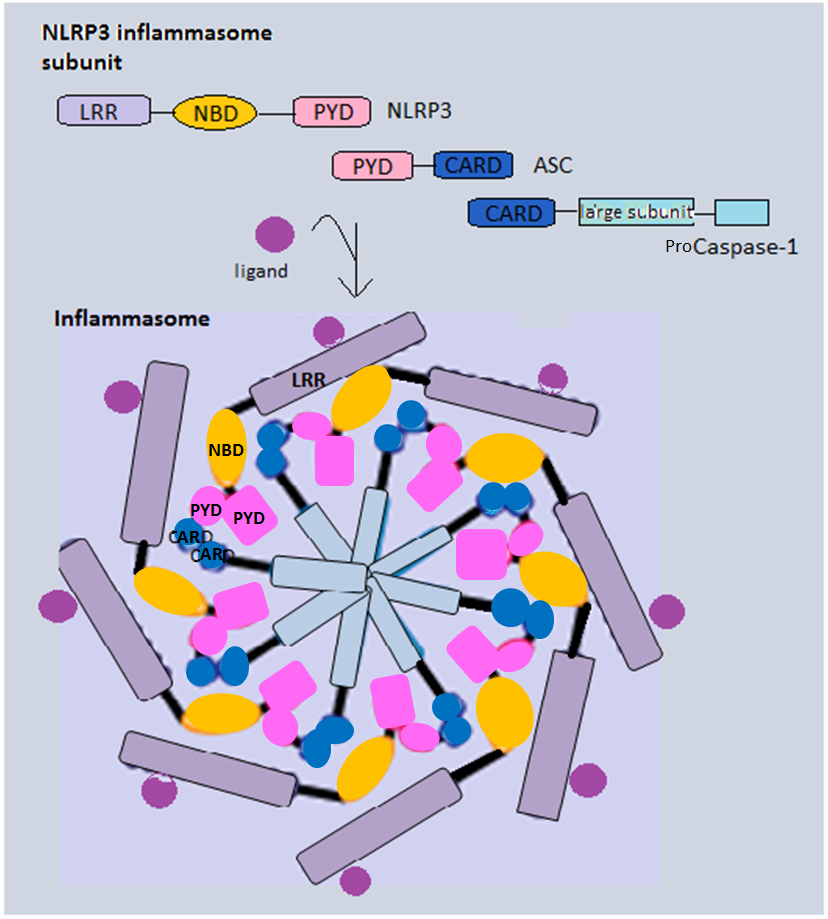
Inflammasomes are cytosolic multiprotein complexes of the innate immune system responsible for the activation of inflammatory responses and cell death. They are formed as a result of specific cytosolic pattern recognition receptors (PRRs) sensing microbe-derived pathogen-associated molecular patterns (PAMPs), damage-associated molecular patterns (DAMPs) from the host cell, or homeostatic disruptions. Activation and assembly of the inflammasome promotes the activation of caspase-1, which then proteolytically cleaves pro-inflammatory cytokines, interleukin 1β (IL-1β) and interleukin 18 (IL-18), as well as the pore-forming molecule gasdermin D (GSDMD). The N-terminal GSDMD fragment resulting from this cleavage induces a pro-inflammatory form of programmed cell death distinct from apoptosis, referred to as pyroptosis, which is responsible for the release of mature cytokines. Additionally, inflammasomes can act as integral components of larger cell death-inducing complexes called PANoptosomes, which drive another distinct form of pro-inflammatory cell death called PANoptosis.
The germline-encoded PRRs that drive inflammasome formation consist of NLRs (nucleotide-binding oligomerization domain and leucine-rich repeat-containing receptors), AIM2 (absent in melanoma 2), IFI16 (IFN-inducible protein 16), and pyrin. Through their caspase activation and recruitment domain (CARD) or pyrin domain (PYD), the inflammasome receptors interact with the adaptor protein called apoptosis-associated speck like protein containing a CARD (also known as ASC or Pycard), which then recruits pro-caspase-1 via its CARD domain to activate inflammatory signaling and pyroptotic cell death. Notably, the PYD of the adaptor protein ASC has been demonstrated to function as a prion-like domain, forming a self-perpetuating polymer when activated. In addition to inflammasomes activating caspase-1, several studies also described non-canonical inflammasome complexes that act independent of caspase-1. In mice, the non-canonical inflammasome is activated by direct sensing of cytosolic bacterial lipopolysaccharide (LPS) by caspase-11, which subsequently induces pyroptotic cell death.[doi:10.1038/nri.2016.58] In human cells, the corresponding caspases of the non-canonical inflammasome are caspase 4 and caspase 5.
Traditionally, inflammasomes have mainly been studied in professional innate immune cells, such as macrophages. However, recent studies indicate high levels of inflammasome component expression in epithelial barrier tissues, where they have been demonstrated to serve as an important first line of defense. Moreover, the dysregulation of inflammasome activation can contribute to the pathology of several major diseases, including cancer, autoimmune disorders, inflammatory conditions, metabolic disorders, and neurodegenerative diseases.
The inflammasome was discovered by the team of Jürg Tschopp, at the University of Lausanne, in 2002. In 2002, it was first reported by Martinon et al. that NLRP1 (NLR family PYD-containing 1) could assemble and oligomerize into a structure in vitro, which activated the caspase-1 cascade, thereby leading to the production of pro-inflammatory cytokines, including IL-1β and IL-18. This NLRP1 multi-molecular complex was dubbed the 'inflammasome', generating significant interest in the following years. During that time, several other inflammasomes were discovered, two of which are also driven by NLRs—NLRP3 (NLR family PYD-containing 3) and NLRC4 (NLR family CARD-containing 4). The physiological relevance of the inflammasome was identified in 2006, when three teams defined the inflammasome's role in diseases such as infection, exposure to toxins, gout, and type 2 diabetes. Several PAMPs and DAMPs, including bacterial RNA and imidazoquinolines, viral DNA, muramyl dipeptide (MDP), asbestos, and silica, were found to induce an inflammasome response. Additional links were also found between metabolic syndrome and NLRP3, one of the inflammasome sensors. These findings paved the way for present-day studies in the fields of innate immunity and cell death, where disease mechanisms and treatments are being investigated in relation to inflammasome assembly and activation.
Inflammasomes can also be formed by PRRs other than NLR proteins. In 2009, Hornung et al. identified a PYHIN (pyrin and HIN domain-containing protein) known as AIM2 that assembles an inflammasome in response to foreign cytoplasmic double-stranded DNA (dsDNA), which several other groups also confirmed in back-to-back studies with Hornung et al. Since then, other non-NLR inflammasome sensors have also been identified.
Assembly of the inflammasome and the resulting inflammatory signaling cascade involve a well-orchestrated mechanism comprising upstream sensors, adapters, and downstream effectors. Inflammasomes play a crucial role in innate immunity via PRR activation in response to an array of stimuli, such as infectious triggers (including bacteria, fungi, viruses, and parasites) and sterile triggers (including ion flux, mitochondrial dysfunction, ROS, and metabolic factors).
Canonical inflammasomes are assembled by the NLRs (including NLRP1, NLRP3, NLRC4), AIM2, and pyrin by recruiting pro-caspase-1 (the precursor molecule of caspase-1), with or without the ASC adaptor.[ In its complete state, the inflammasome appositions together many p45 pro-caspase-1 molecules, inducing their autocatalytic cleavage into p20 and p10 subunits. Caspase-1 then assembles into its active form, which consists of two heterodimers, including a p20 and p10 subunit each.
Caspase-1 activation facilitates the release of the inflammatory cytokines IL-1β and IL-18. The cleavage of GSDMD is also initiated by active caspase-1, which results in pyroptosis (where the cell releases its cytoplasmic content to induce pro-inflammatory signaling). The released IL-1β and IL-18 following inflammasome activation induce interferon gamma (IFN-γ) secretion and natural killer cell activation, inactivation of IL-33, DNA fragmentation and cell pore formation, inhibition of glycolytic enzymes, activation of lipid biosynthesis, and secretion of tissue-repair mediators such as pro-IL-1α. While NLRP1, NLRP3, NLRC4, AIM2, and pyrin are the most well-characterized sensors, IFI16, NLRP6, NLRP7, NLRP9b, NLRP10, NLRP12, and CARD8 also play important roles in inflammasome activation and signaling.
Inflammasome
Inflammasomes are cytosolic multiprotein complexes of the innate immune system responsible for the activation of inflammatory responses and cell death. They are formed as a result of specific cytosolic pattern recognition receptors (PRRs) sensing microbe-derived pathogen-associated molecular patterns (PAMPs), damage-associated molecular patterns (DAMPs) from the host cell, or homeostatic disruptions. Activation and assembly of the inflammasome promotes the activation of caspase-1, which then proteolytically cleaves pro-inflammatory cytokines, interleukin 1β (IL-1β) and interleukin 18 (IL-18), as well as the pore-forming molecule gasdermin D (GSDMD). The N-terminal GSDMD fragment resulting from this cleavage induces a pro-inflammatory form of programmed cell death distinct from apoptosis, referred to as pyroptosis, which is responsible for the release of mature cytokines. Additionally, inflammasomes can act as integral components of larger cell death-inducing complexes called PANoptosomes, which drive another distinct form of pro-inflammatory cell death called PANoptosis.
The germline-encoded PRRs that drive inflammasome formation consist of NLRs (nucleotide-binding oligomerization domain and leucine-rich repeat-containing receptors), AIM2 (absent in melanoma 2), IFI16 (IFN-inducible protein 16), and pyrin. Through their caspase activation and recruitment domain (CARD) or pyrin domain (PYD), the inflammasome receptors interact with the adaptor protein called apoptosis-associated speck like protein containing a CARD (also known as ASC or Pycard), which then recruits pro-caspase-1 via its CARD domain to activate inflammatory signaling and pyroptotic cell death. Notably, the PYD of the adaptor protein ASC has been demonstrated to function as a prion-like domain, forming a self-perpetuating polymer when activated. In addition to inflammasomes activating caspase-1, several studies also described non-canonical inflammasome complexes that act independent of caspase-1. In mice, the non-canonical inflammasome is activated by direct sensing of cytosolic bacterial lipopolysaccharide (LPS) by caspase-11, which subsequently induces pyroptotic cell death.[doi:10.1038/nri.2016.58] In human cells, the corresponding caspases of the non-canonical inflammasome are caspase 4 and caspase 5.
Traditionally, inflammasomes have mainly been studied in professional innate immune cells, such as macrophages. However, recent studies indicate high levels of inflammasome component expression in epithelial barrier tissues, where they have been demonstrated to serve as an important first line of defense. Moreover, the dysregulation of inflammasome activation can contribute to the pathology of several major diseases, including cancer, autoimmune disorders, inflammatory conditions, metabolic disorders, and neurodegenerative diseases.
The inflammasome was discovered by the team of Jürg Tschopp, at the University of Lausanne, in 2002. In 2002, it was first reported by Martinon et al. that NLRP1 (NLR family PYD-containing 1) could assemble and oligomerize into a structure in vitro, which activated the caspase-1 cascade, thereby leading to the production of pro-inflammatory cytokines, including IL-1β and IL-18. This NLRP1 multi-molecular complex was dubbed the 'inflammasome', generating significant interest in the following years. During that time, several other inflammasomes were discovered, two of which are also driven by NLRs—NLRP3 (NLR family PYD-containing 3) and NLRC4 (NLR family CARD-containing 4). The physiological relevance of the inflammasome was identified in 2006, when three teams defined the inflammasome's role in diseases such as infection, exposure to toxins, gout, and type 2 diabetes. Several PAMPs and DAMPs, including bacterial RNA and imidazoquinolines, viral DNA, muramyl dipeptide (MDP), asbestos, and silica, were found to induce an inflammasome response. Additional links were also found between metabolic syndrome and NLRP3, one of the inflammasome sensors. These findings paved the way for present-day studies in the fields of innate immunity and cell death, where disease mechanisms and treatments are being investigated in relation to inflammasome assembly and activation.
Inflammasomes can also be formed by PRRs other than NLR proteins. In 2009, Hornung et al. identified a PYHIN (pyrin and HIN domain-containing protein) known as AIM2 that assembles an inflammasome in response to foreign cytoplasmic double-stranded DNA (dsDNA), which several other groups also confirmed in back-to-back studies with Hornung et al. Since then, other non-NLR inflammasome sensors have also been identified.
Assembly of the inflammasome and the resulting inflammatory signaling cascade involve a well-orchestrated mechanism comprising upstream sensors, adapters, and downstream effectors. Inflammasomes play a crucial role in innate immunity via PRR activation in response to an array of stimuli, such as infectious triggers (including bacteria, fungi, viruses, and parasites) and sterile triggers (including ion flux, mitochondrial dysfunction, ROS, and metabolic factors).
Canonical inflammasomes are assembled by the NLRs (including NLRP1, NLRP3, NLRC4), AIM2, and pyrin by recruiting pro-caspase-1 (the precursor molecule of caspase-1), with or without the ASC adaptor.[ In its complete state, the inflammasome appositions together many p45 pro-caspase-1 molecules, inducing their autocatalytic cleavage into p20 and p10 subunits. Caspase-1 then assembles into its active form, which consists of two heterodimers, including a p20 and p10 subunit each.
Caspase-1 activation facilitates the release of the inflammatory cytokines IL-1β and IL-18. The cleavage of GSDMD is also initiated by active caspase-1, which results in pyroptosis (where the cell releases its cytoplasmic content to induce pro-inflammatory signaling). The released IL-1β and IL-18 following inflammasome activation induce interferon gamma (IFN-γ) secretion and natural killer cell activation, inactivation of IL-33, DNA fragmentation and cell pore formation, inhibition of glycolytic enzymes, activation of lipid biosynthesis, and secretion of tissue-repair mediators such as pro-IL-1α. While NLRP1, NLRP3, NLRC4, AIM2, and pyrin are the most well-characterized sensors, IFI16, NLRP6, NLRP7, NLRP9b, NLRP10, NLRP12, and CARD8 also play important roles in inflammasome activation and signaling.
Recent media
Recent media