Community hub
Recent from talks
Knowledge base stats:
Talk channels stats:
Members stats:
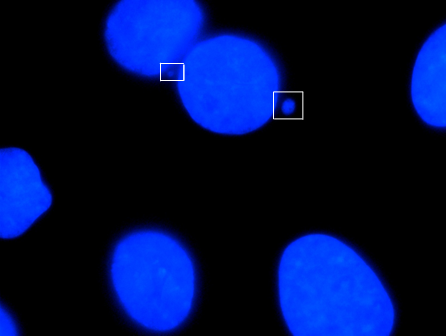
Micronucleus
A micronucleus is a small nucleus that forms whenever a chromosome or a fragment of a chromosome is not incorporated into one of the daughter nuclei during cell division. It usually is a sign of genotoxic events and chromosomal instability. Micronuclei are commonly seen in cancerous cells and may indicate genomic damage events that can increase the risk of developmental or degenerative diseases.
Micronuclei form during anaphase from lagging acentric chromosomes or chromatid fragments caused by incorrectly repaired or unrepaired DNA breaks or by nondisjunction of chromosomes. This improper segregation of chromosomes may result from hypomethylation of repeat sequences present in pericentromeric DNA, irregularities in kinetochore proteins or their assembly, a dysfunctional spindle apparatus, or flawed anaphase checkpoint genes. Micronuclei can contribute to genome instability by promoting a catastrophic mutational event called chromothripsis. Many micronucleus assays have been developed to test for the presence of these structures and determine their frequency in cells exposed to certain chemicals or subjected to stressful conditions.
The term micronucleus may also refer to the smaller nucleus in ciliate protozoans, such as the Paramecium. In mitosis it divides by fission, and in conjugation a pair of gamete micronuclei undergo reciprocal fusion to form a zygote nucleus, which gives rise to the macronuclei and micronuclei of the individuals of the next cycle of fission.
Micronuclei in newly formed red blood cells in humans are known as Howell-Jolly bodies because these structures were first identified and described in erythrocytes by hematologists William Howell and Justin Jolly. These structures were later found to be associated with deficiencies in vitamins such as folate and B12. The relationship between formation of micronuclei and exposure to environmental factors was first reported in root tip cells exposed to ionizing radiation. Micronucleus induction by a chemical was first reported in Ehrlich ascites tumor cells treated with colchicine.
Micronuclei primarily result from acentric chromosome fragments or lagging whole chromosomes that are not included in the daughter nuclei produced by mitosis because they fail to correctly attach to the spindle during the segregation of chromosomes in anaphase. These full chromosomes or chromatid fragments are eventually enclosed by a nuclear membranes and are structurally similar to conventional nuclei, albeit smaller in size. This small nucleus is referred to as a micronucleus. The formation of micronuclei can only be observed in cells undergoing nuclear division and can be clearly seen using cytochalasin B to block cytokinesis to produce a binucleated cells.
Acentric chromosome fragments may arise in a variety of ways. One way is that disrepair of DNA double-strand breaks can lead to symmetrical or asymmetrical chromatid and chromosome exchanges as well as chromatid and chromosome fragments. If DNA damage exceeds the repair capacity of the cell, unrepaired double-stranded DNA breaks may also result in acentric chromosome fragments. Another way eccentric chromosome fragments may arise is when defects in genes related to homologous recombinational repair (ex: ATM, BRCA1, BRCA2, and RAD51) result in a dysfunctional error-free homologous recombinational DNA repair pathway and causes the cell to resort to the error-prone non-homologous end-joining (NHEJ) repair pathway, increasing the likelihood of incorrect repair of DNA breaks, formation of dicentric chromosomes, and acentric chromosome fragments. If enzymes in the NHEJ repair pathway are defective as well, DNA breaks may not be repaired at all. Additionally, simultaneous excision repair of damaged or inappropriate bases incorporated in DNA that are in proximity and on opposite complementary DNA strands may lead to DNA double-stranded breaks and micronucleus formation, especially if the gap-filling step of the repair pathway is not completed.
Micronuclei can also form from fragmented chromosomes when nucleoplasmic bridges (NPB) are formed, stretched, and broken during telophase.
Micronuclei formation may also result from chromosome malsegregation during anaphase. Hypomethylation of cytosine in centromeric and pericentromeric areas and higher-order repeats of satellite DNA in centromeric DNA can result in such chromosomal loss events. Classical satellite DNA is normally heavily methylated at cytosine residues but may become almost fully unmethylated due to ICF syndrome (Immunodeficiency, centromere instability, and facial anomalies syndrome) or after treatment by DNA methyl transferase inhibitors. Since assembly of kinetochore proteins at centromeres is affected by the methylation of cytosine and histone proteins, a reduction in heterochromatin integrity as a result of hypomethylation can interfere with microtubule attachment to chromosomes and with the sensing of tension from correct microtubule-kinetochore connections. Other possible causes of chromosome loss that could lead to micronuclei formation are defects in kinetochore and microtubule interactions, defects in mitotic spindle assembly, mitosis check point defects, abnormal centrosome amplification, and telomeric end fusions that result in dicentric chromosomes that detach from the spindle during anaphase. Micronuclei originating from chromosome loss events and acentric chromosome fragments can be distinguished using pancentromeric DNA probes.
Hub AI
Micronucleus AI simulator
(@Micronucleus_simulator)
Micronucleus
A micronucleus is a small nucleus that forms whenever a chromosome or a fragment of a chromosome is not incorporated into one of the daughter nuclei during cell division. It usually is a sign of genotoxic events and chromosomal instability. Micronuclei are commonly seen in cancerous cells and may indicate genomic damage events that can increase the risk of developmental or degenerative diseases.
Micronuclei form during anaphase from lagging acentric chromosomes or chromatid fragments caused by incorrectly repaired or unrepaired DNA breaks or by nondisjunction of chromosomes. This improper segregation of chromosomes may result from hypomethylation of repeat sequences present in pericentromeric DNA, irregularities in kinetochore proteins or their assembly, a dysfunctional spindle apparatus, or flawed anaphase checkpoint genes. Micronuclei can contribute to genome instability by promoting a catastrophic mutational event called chromothripsis. Many micronucleus assays have been developed to test for the presence of these structures and determine their frequency in cells exposed to certain chemicals or subjected to stressful conditions.
The term micronucleus may also refer to the smaller nucleus in ciliate protozoans, such as the Paramecium. In mitosis it divides by fission, and in conjugation a pair of gamete micronuclei undergo reciprocal fusion to form a zygote nucleus, which gives rise to the macronuclei and micronuclei of the individuals of the next cycle of fission.
Micronuclei in newly formed red blood cells in humans are known as Howell-Jolly bodies because these structures were first identified and described in erythrocytes by hematologists William Howell and Justin Jolly. These structures were later found to be associated with deficiencies in vitamins such as folate and B12. The relationship between formation of micronuclei and exposure to environmental factors was first reported in root tip cells exposed to ionizing radiation. Micronucleus induction by a chemical was first reported in Ehrlich ascites tumor cells treated with colchicine.
Micronuclei primarily result from acentric chromosome fragments or lagging whole chromosomes that are not included in the daughter nuclei produced by mitosis because they fail to correctly attach to the spindle during the segregation of chromosomes in anaphase. These full chromosomes or chromatid fragments are eventually enclosed by a nuclear membranes and are structurally similar to conventional nuclei, albeit smaller in size. This small nucleus is referred to as a micronucleus. The formation of micronuclei can only be observed in cells undergoing nuclear division and can be clearly seen using cytochalasin B to block cytokinesis to produce a binucleated cells.
Acentric chromosome fragments may arise in a variety of ways. One way is that disrepair of DNA double-strand breaks can lead to symmetrical or asymmetrical chromatid and chromosome exchanges as well as chromatid and chromosome fragments. If DNA damage exceeds the repair capacity of the cell, unrepaired double-stranded DNA breaks may also result in acentric chromosome fragments. Another way eccentric chromosome fragments may arise is when defects in genes related to homologous recombinational repair (ex: ATM, BRCA1, BRCA2, and RAD51) result in a dysfunctional error-free homologous recombinational DNA repair pathway and causes the cell to resort to the error-prone non-homologous end-joining (NHEJ) repair pathway, increasing the likelihood of incorrect repair of DNA breaks, formation of dicentric chromosomes, and acentric chromosome fragments. If enzymes in the NHEJ repair pathway are defective as well, DNA breaks may not be repaired at all. Additionally, simultaneous excision repair of damaged or inappropriate bases incorporated in DNA that are in proximity and on opposite complementary DNA strands may lead to DNA double-stranded breaks and micronucleus formation, especially if the gap-filling step of the repair pathway is not completed.
Micronuclei can also form from fragmented chromosomes when nucleoplasmic bridges (NPB) are formed, stretched, and broken during telophase.
Micronuclei formation may also result from chromosome malsegregation during anaphase. Hypomethylation of cytosine in centromeric and pericentromeric areas and higher-order repeats of satellite DNA in centromeric DNA can result in such chromosomal loss events. Classical satellite DNA is normally heavily methylated at cytosine residues but may become almost fully unmethylated due to ICF syndrome (Immunodeficiency, centromere instability, and facial anomalies syndrome) or after treatment by DNA methyl transferase inhibitors. Since assembly of kinetochore proteins at centromeres is affected by the methylation of cytosine and histone proteins, a reduction in heterochromatin integrity as a result of hypomethylation can interfere with microtubule attachment to chromosomes and with the sensing of tension from correct microtubule-kinetochore connections. Other possible causes of chromosome loss that could lead to micronuclei formation are defects in kinetochore and microtubule interactions, defects in mitotic spindle assembly, mitosis check point defects, abnormal centrosome amplification, and telomeric end fusions that result in dicentric chromosomes that detach from the spindle during anaphase. Micronuclei originating from chromosome loss events and acentric chromosome fragments can be distinguished using pancentromeric DNA probes.