Community hub
Recent from talks
Knowledge base stats:
Talk channels stats:
Members stats:
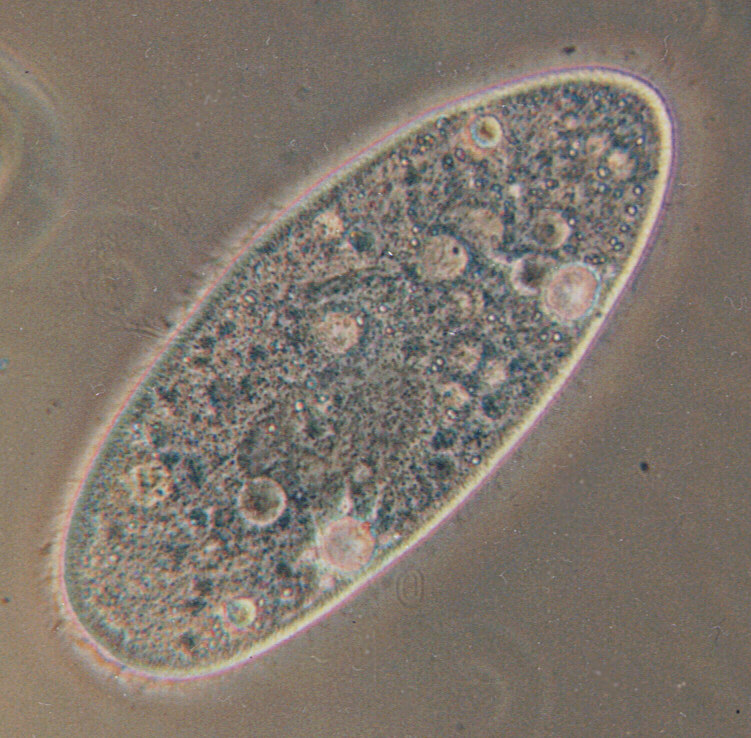
Paramecium
Paramecium (/ˌpærəˈmiːs(i)əm/ PARR-ə-MEE-s(ee-)əm, /-siəm/ -see-əm, plural "paramecia" only when used as a vernacular name) is a genus of eukaryotic, unicellular ciliates, widespread in freshwater, brackish, and marine environments. Paramecia are often abundant in stagnant basins and ponds. Because some species are readily cultivated and easily induced to conjugate and divide, they have been widely used in classrooms and laboratories to study biological processes. Paramecium species are commonly studied as model organisms of the ciliate group and have been characterized as the "white rats" of the phylum Ciliophora.
Paramecium were among the first ciliates to be observed by microscopists, in the late 17th century. They were most likely known to the Dutch pioneer of protozoology, Antonie van Leeuwenhoek, and were clearly described by his contemporary Christiaan Huygens in a letter from 1678. The earliest known illustration of a Paramecium species was published anonymously in Philosophical Transactions of the Royal Society in 1703.
In 1718, the French mathematics teacher and microscopist Louis Joblot published a description and illustration of a microscopic poisson (fish), which he discovered in an infusion of oak bark in water. Joblot gave this creature the name "Chausson", or "slipper", and the phrase "slipper animalcule" remained in use as a colloquial epithet for Paramecium, throughout the 18th and 19th centuries.
The name "Paramecium" – constructed from the Greek παραμήκης (paramēkēs, "oblong") – was coined in 1752 by the English microscopist John Hill, who applied the name generally to "Animalcules which have no visible limbs or tails, and are of an irregularly oblong figure." In 1773, O. F. Müller, the first researcher to place the genus within the Linnaean system of taxonomy, adopted the name Paramecium but changed the spelling to Paramæcium. In 1783, Johann Hermann changed the spelling once more, to Paramœcium. C. G. Ehrenberg, in a major study of the infusoria published in 1838, restored Hill's original spelling for the name, and most researchers have followed his lead.
Species of Paramecium range in size from 0.06 mm to 0.3 mm in length. Cells are typically ovoid, elongate, or foot- or cigar-shaped.
The body of the cell is enclosed by a stiff but elastic structure called the pellicle. The pellicle consists of an outer cell membrane (plasma membrane), a layer of flattened membrane-bound sacs called alveoli, and an inner membrane called the epiplasm. The pellicle is not smooth, but textured with hexagonal or rectangular depressions. Each of these polygons is perforated by a central aperture through which a single cilium projects. Between the alveolar sacs of the pellicle, most species of Paramecium have closely spaced spindle-shaped trichocysts, explosive organelles that discharge thin, non-toxic filaments, often used for defensive purposes.
Typically, an anal pore (cytoproct) is located on the ventral surface, in the posterior half of the cell. In all species, there is a deep oral groove running from the anterior of the cell to its midpoint. This is lined with inconspicuous cilia which beat continuously, drawing food into the cell. Paramecium are primarily heterotrophic, feeding on bacteria and other small organisms. A few species are mixotrophs, deriving some nutrients from endosymbiotic algae (chlorella) carried in the cytoplasm of the cell.
Osmoregulation is carried out by contractile vacuoles, which actively expel water from the cell to compensate for fluid absorbed by osmosis from its surroundings. The number of contractile vacuoles varies depending on the species.
Hub AI
Paramecium AI simulator
(@Paramecium_simulator)
Paramecium
Paramecium (/ˌpærəˈmiːs(i)əm/ PARR-ə-MEE-s(ee-)əm, /-siəm/ -see-əm, plural "paramecia" only when used as a vernacular name) is a genus of eukaryotic, unicellular ciliates, widespread in freshwater, brackish, and marine environments. Paramecia are often abundant in stagnant basins and ponds. Because some species are readily cultivated and easily induced to conjugate and divide, they have been widely used in classrooms and laboratories to study biological processes. Paramecium species are commonly studied as model organisms of the ciliate group and have been characterized as the "white rats" of the phylum Ciliophora.
Paramecium were among the first ciliates to be observed by microscopists, in the late 17th century. They were most likely known to the Dutch pioneer of protozoology, Antonie van Leeuwenhoek, and were clearly described by his contemporary Christiaan Huygens in a letter from 1678. The earliest known illustration of a Paramecium species was published anonymously in Philosophical Transactions of the Royal Society in 1703.
In 1718, the French mathematics teacher and microscopist Louis Joblot published a description and illustration of a microscopic poisson (fish), which he discovered in an infusion of oak bark in water. Joblot gave this creature the name "Chausson", or "slipper", and the phrase "slipper animalcule" remained in use as a colloquial epithet for Paramecium, throughout the 18th and 19th centuries.
The name "Paramecium" – constructed from the Greek παραμήκης (paramēkēs, "oblong") – was coined in 1752 by the English microscopist John Hill, who applied the name generally to "Animalcules which have no visible limbs or tails, and are of an irregularly oblong figure." In 1773, O. F. Müller, the first researcher to place the genus within the Linnaean system of taxonomy, adopted the name Paramecium but changed the spelling to Paramæcium. In 1783, Johann Hermann changed the spelling once more, to Paramœcium. C. G. Ehrenberg, in a major study of the infusoria published in 1838, restored Hill's original spelling for the name, and most researchers have followed his lead.
Species of Paramecium range in size from 0.06 mm to 0.3 mm in length. Cells are typically ovoid, elongate, or foot- or cigar-shaped.
The body of the cell is enclosed by a stiff but elastic structure called the pellicle. The pellicle consists of an outer cell membrane (plasma membrane), a layer of flattened membrane-bound sacs called alveoli, and an inner membrane called the epiplasm. The pellicle is not smooth, but textured with hexagonal or rectangular depressions. Each of these polygons is perforated by a central aperture through which a single cilium projects. Between the alveolar sacs of the pellicle, most species of Paramecium have closely spaced spindle-shaped trichocysts, explosive organelles that discharge thin, non-toxic filaments, often used for defensive purposes.
Typically, an anal pore (cytoproct) is located on the ventral surface, in the posterior half of the cell. In all species, there is a deep oral groove running from the anterior of the cell to its midpoint. This is lined with inconspicuous cilia which beat continuously, drawing food into the cell. Paramecium are primarily heterotrophic, feeding on bacteria and other small organisms. A few species are mixotrophs, deriving some nutrients from endosymbiotic algae (chlorella) carried in the cytoplasm of the cell.
Osmoregulation is carried out by contractile vacuoles, which actively expel water from the cell to compensate for fluid absorbed by osmosis from its surroundings. The number of contractile vacuoles varies depending on the species.