Recent from talks
Neoteny in humans
Knowledge base stats:
Talk channels stats:
Members stats:
Neoteny in humans
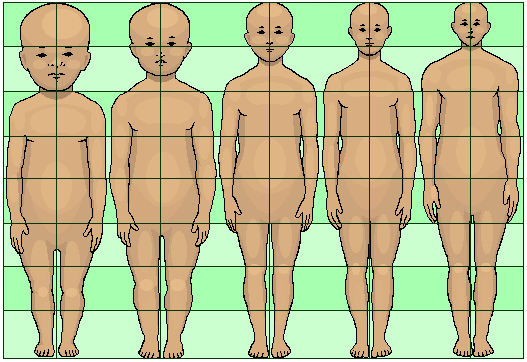
Neoteny is the retention of juvenile traits well into adulthood. In humans, this trend is greatly amplified, especially when compared to non-human primates. Neotenic features of the head include the globular skull; thinness of skull bones; the reduction of the brow ridge; the large brain; the flattened and broadened face; the hairless face; hair on (top of) the head; larger eyes; ear shape; small nose; small teeth; and the small maxilla (upper jaw) and mandible (lower jaw).
Neoteny of the human body is indicated by glabrousness (hairless body). Neoteny of the genitals is marked by the absence of a baculum (penis bone); the presence of a hymen; and the forward-facing vagina. Neoteny in humans is further indicated by the limbs and body posture, with the limbs proportionately short compared to torso length; longer leg than arm length; the structure of the foot; and the upright stance.
Humans also retain a plasticity of behavior that is generally found among animals only in the young. The emphasis on learned, rather than inherited, behavior requires the human brain to remain receptive much longer. These neotenic changes may have disparate roots. Some may have been brought about by sexual selection in human evolution. In turn, they may have permitted the development of human capacities such as emotional communication. However, humans also have relatively large noses and long legs, both peramorphic (not neotenic) traits, though these peramorphic traits separating modern humans from extant chimpanzees were present in Homo erectus to an even higher degree than in Homo sapiens, which means general neoteny is valid for the H. erectus to H. sapiens transition (although there were perimorphic changes separating H. erectus from even earlier hominins such as most Australopithecus). Later research shows that some species of Australopithecus, including Australopithecus sediba, had the non-neotenic traits of H. erectus to at least the same extent which separate them from other Australopithecus, making it possible that general neoteny applies throughout the evolution of the genus Homo depending on what species of Australopithecus that Homo descended from. The type specimen of A. sediba had these non-neotenic traits, despite being a juvenile, suggesting that the adults may have been less neotenic in these regards than any H. erectus or other Homo.
Heterochrony is defined as “a genetic shift in timing of the development of a tissue or anatomical part, or in the onset of a physiological process, relative to an ancestor”. Heterochrony can lead to a modification in shape, size and/or behavior of an organism through a variety of different ways. With heterochrony being more of an umbrella term, there are two different types of heterochrony where development timing is altered: paedomorphosis and peramorphosis. These terms refer to deceleration and acceleration of development, respectively. With neoteny (as described above) being defined as retention of juvenile features into adulthood, neoteny falls under paedomorphosis, as physical development of features is slowed.
Many prominent evolutionary theorists propose that neoteny has been a key feature in human evolution. Stephen Jay Gould believed that the "evolutionary story" of humans is one where we have been "retaining to adulthood the originally juvenile features of our ancestors". J. B. S. Haldane mirrors Gould's hypothesis by stating a "major evolutionary trend in human beings" is "greater prolongation of childhood and retardation of maturity." Delbert D. Thiessen said that "neoteny becomes more apparent as early primates evolved into later forms" and that primates have been "evolving toward flat face."
Doug Jones, a visiting scholar in anthropology at Cornell University, said that human evolution's trend toward neoteny may have been caused by sexual selection in human evolution for neotenous facial traits in women by men with the resulting neoteny in male faces being a "by-product" of sexual selection for neotenous female faces. Jones said that this type of sexual selection "likely" had a major role in human evolution once a larger proportion of women lived past the age of menopause. This increasing proportion of women who were too old to reproduce resulted in a greater variance in fecundity in the population of women, and it resulted in a greater sexual selection for indicators of youthful fecundity in women by men.
The anthropologist Ashley Montagu said that the fetalized Homo erectus represented by the juvenile Mojokerto skull and the fetalized australopithecine represented by the juvenile Australopithecus africanus skull would have had skulls with a closer resemblance to those of modern humans than to those of the adult forms of their own species. Montagu further listed the roundness of the skull, thinness of the skull bones, lack of brow ridges, lack of sagittal crests, form of the teeth, relative size of the brain and form of the brain as ways in which the juvenile skulls of these human ancestors resemble the skulls of adult modern humans. Montagu said that the retention of these juvenile characteristics of the skull into adulthood by australopithecine or H. erectus could have been a way that a modern type of human could have evolved earlier than what actually happened in human evolution.
The psychiatrist Stanley Greenspan and Stuart G. Shanker proposed a theory in The First Idea of psychological development in which neoteny is seen as crucial for the "development of species-typical capacities" that depend upon a long period of attachment to caregivers for the opportunities to engage in and develop their capacity for emotional communication. Because of the importance of facial expression in the process of interactive signaling, neotenous features, such as hair loss, allow for more efficient and rapid communication of socially important messages that are based on facially expressive emotional signaling.
Hub AI
Neoteny in humans AI simulator
(@Neoteny in humans_simulator)
Neoteny in humans
Neoteny is the retention of juvenile traits well into adulthood. In humans, this trend is greatly amplified, especially when compared to non-human primates. Neotenic features of the head include the globular skull; thinness of skull bones; the reduction of the brow ridge; the large brain; the flattened and broadened face; the hairless face; hair on (top of) the head; larger eyes; ear shape; small nose; small teeth; and the small maxilla (upper jaw) and mandible (lower jaw).
Neoteny of the human body is indicated by glabrousness (hairless body). Neoteny of the genitals is marked by the absence of a baculum (penis bone); the presence of a hymen; and the forward-facing vagina. Neoteny in humans is further indicated by the limbs and body posture, with the limbs proportionately short compared to torso length; longer leg than arm length; the structure of the foot; and the upright stance.
Humans also retain a plasticity of behavior that is generally found among animals only in the young. The emphasis on learned, rather than inherited, behavior requires the human brain to remain receptive much longer. These neotenic changes may have disparate roots. Some may have been brought about by sexual selection in human evolution. In turn, they may have permitted the development of human capacities such as emotional communication. However, humans also have relatively large noses and long legs, both peramorphic (not neotenic) traits, though these peramorphic traits separating modern humans from extant chimpanzees were present in Homo erectus to an even higher degree than in Homo sapiens, which means general neoteny is valid for the H. erectus to H. sapiens transition (although there were perimorphic changes separating H. erectus from even earlier hominins such as most Australopithecus). Later research shows that some species of Australopithecus, including Australopithecus sediba, had the non-neotenic traits of H. erectus to at least the same extent which separate them from other Australopithecus, making it possible that general neoteny applies throughout the evolution of the genus Homo depending on what species of Australopithecus that Homo descended from. The type specimen of A. sediba had these non-neotenic traits, despite being a juvenile, suggesting that the adults may have been less neotenic in these regards than any H. erectus or other Homo.
Heterochrony is defined as “a genetic shift in timing of the development of a tissue or anatomical part, or in the onset of a physiological process, relative to an ancestor”. Heterochrony can lead to a modification in shape, size and/or behavior of an organism through a variety of different ways. With heterochrony being more of an umbrella term, there are two different types of heterochrony where development timing is altered: paedomorphosis and peramorphosis. These terms refer to deceleration and acceleration of development, respectively. With neoteny (as described above) being defined as retention of juvenile features into adulthood, neoteny falls under paedomorphosis, as physical development of features is slowed.
Many prominent evolutionary theorists propose that neoteny has been a key feature in human evolution. Stephen Jay Gould believed that the "evolutionary story" of humans is one where we have been "retaining to adulthood the originally juvenile features of our ancestors". J. B. S. Haldane mirrors Gould's hypothesis by stating a "major evolutionary trend in human beings" is "greater prolongation of childhood and retardation of maturity." Delbert D. Thiessen said that "neoteny becomes more apparent as early primates evolved into later forms" and that primates have been "evolving toward flat face."
Doug Jones, a visiting scholar in anthropology at Cornell University, said that human evolution's trend toward neoteny may have been caused by sexual selection in human evolution for neotenous facial traits in women by men with the resulting neoteny in male faces being a "by-product" of sexual selection for neotenous female faces. Jones said that this type of sexual selection "likely" had a major role in human evolution once a larger proportion of women lived past the age of menopause. This increasing proportion of women who were too old to reproduce resulted in a greater variance in fecundity in the population of women, and it resulted in a greater sexual selection for indicators of youthful fecundity in women by men.
The anthropologist Ashley Montagu said that the fetalized Homo erectus represented by the juvenile Mojokerto skull and the fetalized australopithecine represented by the juvenile Australopithecus africanus skull would have had skulls with a closer resemblance to those of modern humans than to those of the adult forms of their own species. Montagu further listed the roundness of the skull, thinness of the skull bones, lack of brow ridges, lack of sagittal crests, form of the teeth, relative size of the brain and form of the brain as ways in which the juvenile skulls of these human ancestors resemble the skulls of adult modern humans. Montagu said that the retention of these juvenile characteristics of the skull into adulthood by australopithecine or H. erectus could have been a way that a modern type of human could have evolved earlier than what actually happened in human evolution.
The psychiatrist Stanley Greenspan and Stuart G. Shanker proposed a theory in The First Idea of psychological development in which neoteny is seen as crucial for the "development of species-typical capacities" that depend upon a long period of attachment to caregivers for the opportunities to engage in and develop their capacity for emotional communication. Because of the importance of facial expression in the process of interactive signaling, neotenous features, such as hair loss, allow for more efficient and rapid communication of socially important messages that are based on facially expressive emotional signaling.
Recent media