Recent from talks
Phosphofructokinase 2
Knowledge base stats:
Talk channels stats:
Members stats:
Phosphofructokinase 2
Phosphofructokinase-2 (6-phosphofructo-2-kinase, PFK-2) or fructose bisphosphatase-2 (FBPase-2), is an enzyme indirectly responsible for regulating the rates of glycolysis and gluconeogenesis in cells. It catalyzes formation and degradation of a significant allosteric regulator, fructose-2,6-bisphosphate (Fru-2,6-P2) from substrate fructose-6-phosphate. Fru-2,6-P2 contributes to the rate-determining step of glycolysis as it activates enzyme phosphofructokinase 1 in the glycolysis pathway, and inhibits fructose-1,6-bisphosphatase 1 in gluconeogenesis. Since Fru-2,6-P2 differentially regulates glycolysis and gluconeogenesis, it can act as a key signal to switch between the opposing pathways. Because PFK-2 produces Fru-2,6-P2 in response to hormonal signaling, metabolism can be more sensitively and efficiently controlled to align with the organism's glycolytic needs. This enzyme participates in fructose and mannose metabolism. The enzyme is important in the regulation of hepatic carbohydrate metabolism and is found in greatest quantities in the liver, kidney and heart. In mammals, several genes often encode different isoforms, each of which differs in its tissue distribution and enzymatic activity. The family described here bears a resemblance to the ATP-driven phospho-fructokinases; however, they share little sequence similarity, although a few residues seem key to their interaction with fructose 6-phosphate.
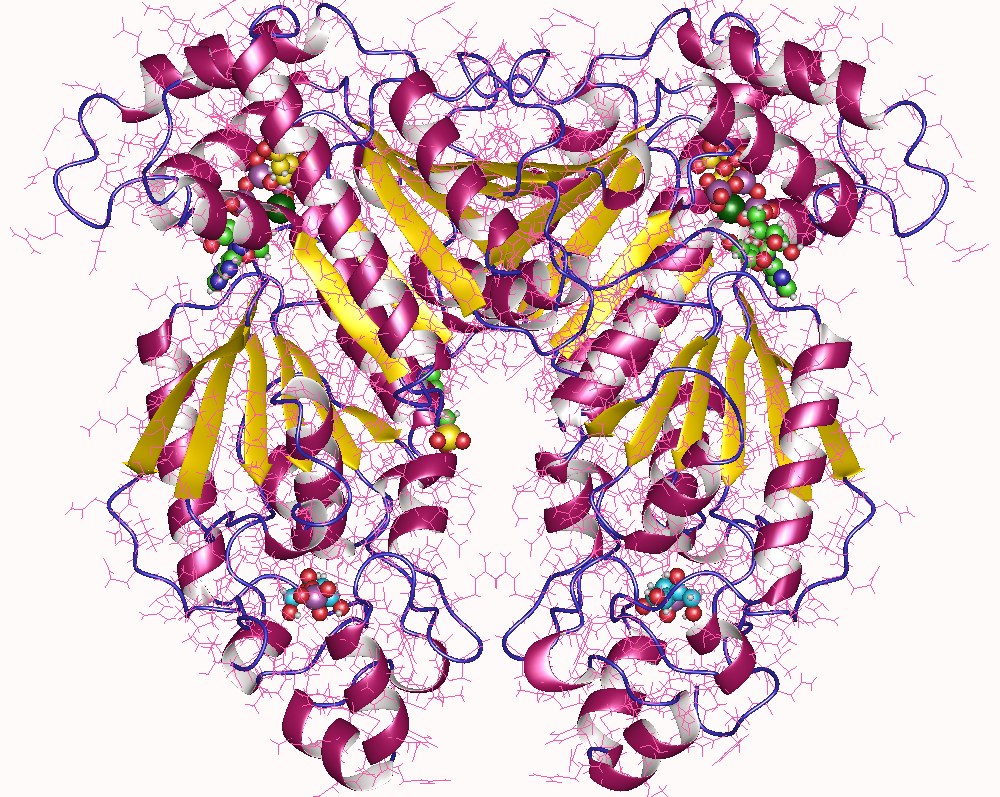
PFK-2 is known as the "bifunctional enzyme" because of its notable structure: though both are located on one protein homodimer, its two domains act as independently functioning enzymes. One terminus serves as a kinase domain (for PFK-2) while the other terminus acts as a phosphatase domain (FBPase-2).
In mammals, genetic mechanisms encode different PFK-2 isoforms to accommodate tissue specific needs. While general function remains the same, isoforms feature slight differences in enzymatic properties and are controlled by different methods of regulation; these differences are discussed below.
The monomers of the bifunctional protein are clearly divided into two functional domains. The kinase domain is located on the N-terminal. It consists of a central six-stranded β sheet, with five parallel strands and an antiparallel edge strand, surrounded by seven α helices. The domain contains nucleotide-binding fold (nbf) at the C-terminal end of the first β-strand. The PFK-2 domain appears to be closely related to the superfamily of mononucleotide binding proteins including adenylate cyclase.
On the other hand, the phosphatase domain is located on the C-terminal. It resembles the family of proteins that include phosphoglycerate mutases and acid phosphatases. The domain has a mixed α/ β structure, with a six-stranded central β sheet, plus an additional α-helical subdomain that covers the presumed active site of the molecule. Finally, the N-terminal region modulates PFK-2 and FBPase2 activities, and stabilizes the dimer form of the enzyme.
While this central catalytic core remains conserved in all forms of PFK-2, slight structural variations exist in isoforms as a result of different amino acid sequences or alternative splicing. With some minor exceptions, the size of PFK-2 enzymes is typically around 55 kDa.
Researchers hypothesize that the unique bifunctional structure of this enzyme arose from a gene fusion event between a primordial bacterial PFK-1 and a primordial mutase/phosphatase.
This enzyme's main function is to synthesize or degrade allosteric regulator Fru-2,6-P2 in response to glycolytic needs of the cell or organism, as depicted in the accompanying diagram.
Hub AI
Phosphofructokinase 2 AI simulator
(@Phosphofructokinase 2_simulator)
Phosphofructokinase 2
Phosphofructokinase-2 (6-phosphofructo-2-kinase, PFK-2) or fructose bisphosphatase-2 (FBPase-2), is an enzyme indirectly responsible for regulating the rates of glycolysis and gluconeogenesis in cells. It catalyzes formation and degradation of a significant allosteric regulator, fructose-2,6-bisphosphate (Fru-2,6-P2) from substrate fructose-6-phosphate. Fru-2,6-P2 contributes to the rate-determining step of glycolysis as it activates enzyme phosphofructokinase 1 in the glycolysis pathway, and inhibits fructose-1,6-bisphosphatase 1 in gluconeogenesis. Since Fru-2,6-P2 differentially regulates glycolysis and gluconeogenesis, it can act as a key signal to switch between the opposing pathways. Because PFK-2 produces Fru-2,6-P2 in response to hormonal signaling, metabolism can be more sensitively and efficiently controlled to align with the organism's glycolytic needs. This enzyme participates in fructose and mannose metabolism. The enzyme is important in the regulation of hepatic carbohydrate metabolism and is found in greatest quantities in the liver, kidney and heart. In mammals, several genes often encode different isoforms, each of which differs in its tissue distribution and enzymatic activity. The family described here bears a resemblance to the ATP-driven phospho-fructokinases; however, they share little sequence similarity, although a few residues seem key to their interaction with fructose 6-phosphate.
PFK-2 is known as the "bifunctional enzyme" because of its notable structure: though both are located on one protein homodimer, its two domains act as independently functioning enzymes. One terminus serves as a kinase domain (for PFK-2) while the other terminus acts as a phosphatase domain (FBPase-2).
In mammals, genetic mechanisms encode different PFK-2 isoforms to accommodate tissue specific needs. While general function remains the same, isoforms feature slight differences in enzymatic properties and are controlled by different methods of regulation; these differences are discussed below.
The monomers of the bifunctional protein are clearly divided into two functional domains. The kinase domain is located on the N-terminal. It consists of a central six-stranded β sheet, with five parallel strands and an antiparallel edge strand, surrounded by seven α helices. The domain contains nucleotide-binding fold (nbf) at the C-terminal end of the first β-strand. The PFK-2 domain appears to be closely related to the superfamily of mononucleotide binding proteins including adenylate cyclase.
On the other hand, the phosphatase domain is located on the C-terminal. It resembles the family of proteins that include phosphoglycerate mutases and acid phosphatases. The domain has a mixed α/ β structure, with a six-stranded central β sheet, plus an additional α-helical subdomain that covers the presumed active site of the molecule. Finally, the N-terminal region modulates PFK-2 and FBPase2 activities, and stabilizes the dimer form of the enzyme.
While this central catalytic core remains conserved in all forms of PFK-2, slight structural variations exist in isoforms as a result of different amino acid sequences or alternative splicing. With some minor exceptions, the size of PFK-2 enzymes is typically around 55 kDa.
Researchers hypothesize that the unique bifunctional structure of this enzyme arose from a gene fusion event between a primordial bacterial PFK-1 and a primordial mutase/phosphatase.
This enzyme's main function is to synthesize or degrade allosteric regulator Fru-2,6-P2 in response to glycolytic needs of the cell or organism, as depicted in the accompanying diagram.
Recent media