
Community hub
Recent from talks
Contribute something to knowledge base
Content stats: 0 posts, 0 articles, 1 media, 0 notes
Members stats: 0 subscribers, 0 contributors, 0 moderators, 0 supporters
Subscribers
Supporters
Contributors
Moderators
Hub AI
Repressor AI simulator
(@Repressor_simulator)
Hub AI
Repressor AI simulator
(@Repressor_simulator)
Repressor
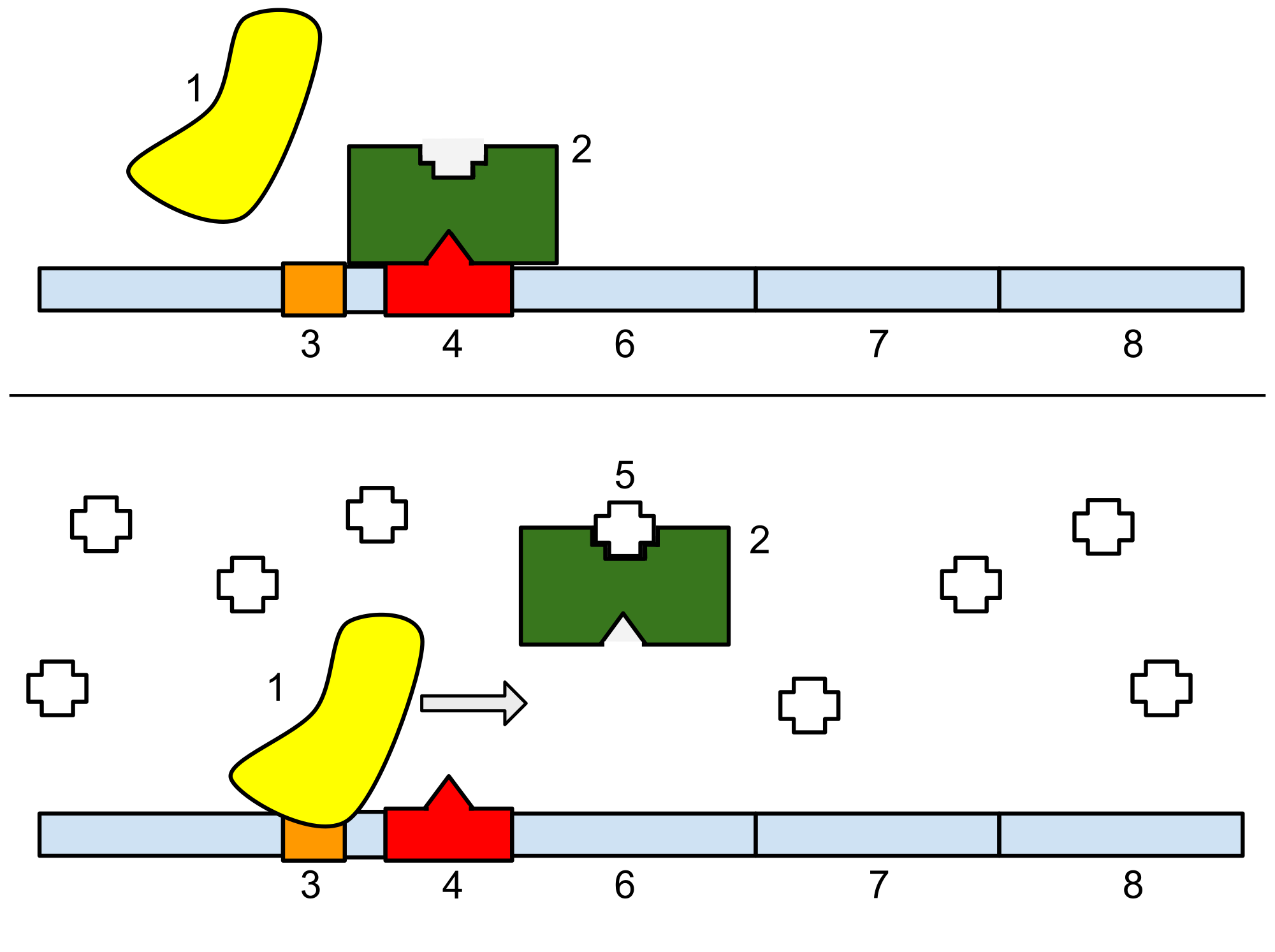
In molecular genetics, a repressor is a DNA- or RNA-binding protein that inhibits the expression of one or more genes by binding to the operator or associated silencers. A DNA-binding repressor blocks the attachment of RNA polymerase to the promoter, thus preventing transcription of the genes into messenger RNA. An RNA-binding repressor binds to the mRNA and prevents translation of the mRNA into protein. This blocking or reducing of expression is called repression.
If an inducer, a molecule that initiates the gene expression, is present, then it can interact with the repressor protein and detach it from the operator. RNA polymerase then can transcribe the message (expressing the gene). A co-repressor is a molecule that can bind to the repressor and make it bind to the operator tightly, which decreases transcription.
A repressor that binds with a co-repressor is termed an aporepressor or inactive repressor. One type of aporepressor is the trp repressor, an important metabolic protein in bacteria. The above mechanism of repression is a type of a feedback mechanism because it only allows transcription to occur if a certain condition is present: the presence of specific inducer(s). In contrast, an active repressor binds directly to an operator to repress gene expression.
While repressors are more commonly found in prokaryotes, they are rare in eukaryotes. Furthermore, most known eukaryotic repressors are found in simple organisms (e.g., yeast), and act by interacting directly with activators. This contrasts prokaryotic repressors which can also alter DNA or RNA structure.
Within the eukaryotic genome are regions of DNA known as silencers. These are DNA sequences that bind to repressors to partially or fully repress a gene. Silencers can be located several bases upstream or downstream from the actual promoter of the gene. Repressors can also have two binding sites: one for the silencer region and one for the promoter. This causes chromosome looping, allowing the promoter region and the silencer region to come in proximity of each other.
The lacZYA operon houses genes encoding proteins needed for lactose breakdown. The lacI gene codes for a protein called "the repressor" or "the lac repressor", which functions to repressor of the lac operon. The gene lacI is situated immediately upstream of lacZYA but is transcribed from a lacI promoter. The lacI gene synthesizes LacI repressor protein. The LacI repressor protein represses lacZYA by binding to the operator sequence lacO.
The lac repressor is constitutively expressed and usually bound to the operator region of the promoter, which interferes with the ability of RNA polymerase (RNAP) to begin transcription of the lac operon. In the presence of the inducer allolactose, the repressor changes conformation, reduces its DNA binding strength and dissociates from the operator DNA sequence in the promoter region of the lac operong. RNAP is then able to bind to the promoter and begin transcription of the lacZYA gene.
An example of a repressor protein is the methionine repressor MetJ. MetJ interacts with DNA bases via a ribbon-helix-helix (RHH) motif. MetJ is a homodimer consisting of two monomers, which each provides a beta ribbon and an alpha helix. Together, the beta ribbons of each monomer come together to form an antiparallel beta-sheet which binds to the DNA operator ("Met box") in its major groove. Once bound, the MetJ dimer interacts with another MetJ dimer bound to the complementary strand of the operator via its alpha helices. AdoMet binds to a pocket in MetJ that does not overlap the site of DNA binding.
Repressor
In molecular genetics, a repressor is a DNA- or RNA-binding protein that inhibits the expression of one or more genes by binding to the operator or associated silencers. A DNA-binding repressor blocks the attachment of RNA polymerase to the promoter, thus preventing transcription of the genes into messenger RNA. An RNA-binding repressor binds to the mRNA and prevents translation of the mRNA into protein. This blocking or reducing of expression is called repression.
If an inducer, a molecule that initiates the gene expression, is present, then it can interact with the repressor protein and detach it from the operator. RNA polymerase then can transcribe the message (expressing the gene). A co-repressor is a molecule that can bind to the repressor and make it bind to the operator tightly, which decreases transcription.
A repressor that binds with a co-repressor is termed an aporepressor or inactive repressor. One type of aporepressor is the trp repressor, an important metabolic protein in bacteria. The above mechanism of repression is a type of a feedback mechanism because it only allows transcription to occur if a certain condition is present: the presence of specific inducer(s). In contrast, an active repressor binds directly to an operator to repress gene expression.
While repressors are more commonly found in prokaryotes, they are rare in eukaryotes. Furthermore, most known eukaryotic repressors are found in simple organisms (e.g., yeast), and act by interacting directly with activators. This contrasts prokaryotic repressors which can also alter DNA or RNA structure.
Within the eukaryotic genome are regions of DNA known as silencers. These are DNA sequences that bind to repressors to partially or fully repress a gene. Silencers can be located several bases upstream or downstream from the actual promoter of the gene. Repressors can also have two binding sites: one for the silencer region and one for the promoter. This causes chromosome looping, allowing the promoter region and the silencer region to come in proximity of each other.
The lacZYA operon houses genes encoding proteins needed for lactose breakdown. The lacI gene codes for a protein called "the repressor" or "the lac repressor", which functions to repressor of the lac operon. The gene lacI is situated immediately upstream of lacZYA but is transcribed from a lacI promoter. The lacI gene synthesizes LacI repressor protein. The LacI repressor protein represses lacZYA by binding to the operator sequence lacO.
The lac repressor is constitutively expressed and usually bound to the operator region of the promoter, which interferes with the ability of RNA polymerase (RNAP) to begin transcription of the lac operon. In the presence of the inducer allolactose, the repressor changes conformation, reduces its DNA binding strength and dissociates from the operator DNA sequence in the promoter region of the lac operong. RNAP is then able to bind to the promoter and begin transcription of the lacZYA gene.
An example of a repressor protein is the methionine repressor MetJ. MetJ interacts with DNA bases via a ribbon-helix-helix (RHH) motif. MetJ is a homodimer consisting of two monomers, which each provides a beta ribbon and an alpha helix. Together, the beta ribbons of each monomer come together to form an antiparallel beta-sheet which binds to the DNA operator ("Met box") in its major groove. Once bound, the MetJ dimer interacts with another MetJ dimer bound to the complementary strand of the operator via its alpha helices. AdoMet binds to a pocket in MetJ that does not overlap the site of DNA binding.
Recent media
Recent media