Community hub
0 subscribers8 pages, 0 posts
Recent from talks
All channels
Be the first to start a discussion here.
Be the first to start a discussion here.
Be the first to start a discussion here.
Be the first to start a discussion here.
Contribute something
Welcome to the community hub built to collect knowledge and have discussions related to Statocyst.
Nothing was collected or created yet.
Statocyst
View on Wikipediafrom Wikipedia
Not found
Statocyst
View on Grokipediafrom Grokipedia
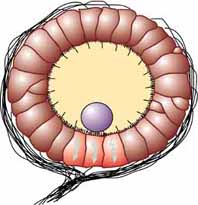
A statocyst is a specialized sensory organ found in many aquatic invertebrates, functioning as a gravity receptor to detect linear and rotational accelerations for maintaining equilibrium and orientation in three-dimensional space.[1] It typically consists of a fluid-filled, closed sac lined with ciliated mechanoreceptor hair cells and containing dense mineral particles known as statoliths or statoconia, which lag behind during movement and deflect the sensory hairs to transduce gravitational or acceleratory stimuli into neural signals.[1] These organs are analogous—and likely homologous—to the otolith organs of the vertebrate inner ear, enabling precise detection of body position despite the challenges of buoyancy and viscosity in aquatic environments.[1]
Statocysts occur across a wide range of invertebrate phyla, including Cnidaria, Ctenophora, Platyhelminthes, Mollusca, Annelida, Arthropoda, and Echinodermata, reflecting their ancient evolutionary origin predating the divergence of Bilateria.[1] In simpler forms, such as hydromedusae or flatworms, they may feature a single statolith resting on a few sensory cells, while more complex variants exhibit structural elaborations like multiple sensory fields or toroidal arrangements.[1] For instance, in crustaceans like the Norwegian lobster (Nephrops norvegicus), the statocyst is an invaginated sac in the antennule base, housing arrays of sensory setae embedded with statoconia in a gelatinous matrix to sense both gravity and sound particle motion for behaviors such as locomotion and posture regulation.[2] In cephalopods, such as the nautilus (Nautilus pompilius), statocysts are ovoid chambers lined with 130,000 to 150,000 polarized hair cells of two morphological types, capable of detecting angular accelerations without additional crista/cupula systems found in more derived species like squid.[3]
Beyond geotactic orientation—such as righting responses or navigation to food sources—statocysts play critical roles in vital physiological processes, including sound perception in marine species and compensatory movements during swimming or burrowing.[2] Their sensitivity to mechanical disturbances underscores their vulnerability to anthropogenic noise pollution, which can cause structural damage and impair sensory function, as observed in decapod crustaceans exposed to seismic air guns.[4] Overall, these organs exemplify convergent evolution in sensory solutions for gravity sensing, supporting diverse invertebrate lifestyles from pelagic drifting to benthic foraging.[1]