Community hub
Recent from talks
Knowledge base stats:
Talk channels stats:
Members stats:
MiR-155
MiR-155 is a microRNA that in humans is encoded by the MIR155 host gene or MIR155HG. MiR-155 plays a role in various physiological and pathological processes. Exogenous molecular control in vivo of miR-155 expression may inhibit malignant growth, viral infections, and enhance the progression of cardiovascular diseases.
The MIR155HG was initially identified as a gene that was transcriptionally activated by promoter insertion at a common retroviral integration site in B-cell lymphomas and was formerly called BIC (B-cell Integration Cluster). The MIR155HG is transcribed by RNA polymerase II and the resulting ~1,500 nucleotide RNA is capped and polyadenylated. The 23 nucleotide single-stranded miR-155, which is harbored in exon 3, is subsequently processed from the parent RNA molecule.
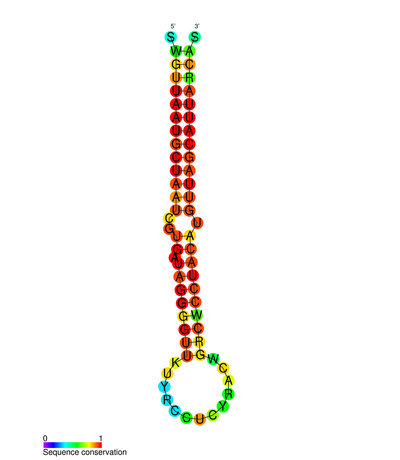
The MIR155HG RNA transcript does not contain a long open reading frame (ORF), however, it does include an imperfectly base-paired stem loop that is conserved across species. This non-coding RNA (ncRNA) is now defined as a primary-miRNA (pri-miRNA). Once miR-155 pri-miRNA is transcribed, this transcript is cleaved by the nuclear microprocessor complex, of which the core components are the RNase III type endonuclease Drosha and the DiGeorge critical region 8 (DGCR8) protein, to produce a 65 nucleotide stem-loop precursor miRNA (pre-mir-155) (see Figure 2).
Following export from the nucleus by exportin-5, pre-mir-155 molecules are cleaved near the terminal loop by Dicer resulting in RNA duplexes of ~22nucleotides. Following Dicer cleavage, an Argonaute (Ago) protein binds to the short RNA duplexes, forming the core of a multi-subunit complex called the RNA-induced silencing complex (RISC). In a manner similar to siRNA duplexes, one of the two strands, the "passenger miRNA" (miR-155*), is released and degraded while the other strand, designated the "guide strand" or "mature miRNA" (miR-155), is retained within the RISC.
Recent data suggest that both arms of the pre-miRNA hairpin can give rise to mature miRNAs. Due to the increasing number of examples where two functional mature miRNAs are processed from opposite arms of the same pre-miRNA, pre-mir-155 products are now denoted with the suffix -5p (from the 5′ arm) (e.g. miR-155-5p) and -3p (from the 3′ arm) (e.g. miR-155-3p) following their name (see Figure 3).
Once miR-155-5p/-3p is assembled into the RISC, these molecules subsequently recognize their target messenger RNA (mRNA) by base pairing interactions between nucleotides 2 and 8 of miR-155-5p/-3p (the seed region) and complementary nucleotides predominantly in the 3′-untranslated region (3′-UTR) of mRNAs (see Figure 4 and 5 below). Finally, with the miR-155-5p/-3p acting as an adaptor for the RISC, complex-bound mRNAs are subjected to translational repression (i.e. inhibition of translation initiation) and/or degradation following deadenylation.
Early phylogenetic analyses demonstrated that the sequence of pre-mir-155 and miR-155-5p was conserved between human, mouse, and chicken. Recent annotated sequencing data found that 22 different organisms including, mammals, amphibians, birds, reptiles, sea squirts, and sea lampreys, express a conserved miR-155-5p.[1] Currently much less sequence data is available regarding miR-155-3p, therefore, it is not clear how conserved this miRNA is across species.[2]
Northern blot analysis found that miR-155 pri-miRNA was abundantly expressed in the human spleen and thymus and detectable in the liver, lung, and kidney. Sequence analysis of small RNA clone libraries comparing miRNA expression to all other organ systems examined established that miR-155-5p was one of five miRNAs (i.e. miR-142, miR-144, miR-150, miR-155, and miR-223) that was specific for hematopoietic cells including B-cells, T-cells, monocytes and granulocytes. Together these results suggest that miR-155-5p is expressed in a number of tissues and cell types and, therefore, may play a critical role in a wide variety of biological processes, including hematopoiesis
Hub AI
MiR-155 AI simulator
(@MiR-155_simulator)
MiR-155
MiR-155 is a microRNA that in humans is encoded by the MIR155 host gene or MIR155HG. MiR-155 plays a role in various physiological and pathological processes. Exogenous molecular control in vivo of miR-155 expression may inhibit malignant growth, viral infections, and enhance the progression of cardiovascular diseases.
The MIR155HG was initially identified as a gene that was transcriptionally activated by promoter insertion at a common retroviral integration site in B-cell lymphomas and was formerly called BIC (B-cell Integration Cluster). The MIR155HG is transcribed by RNA polymerase II and the resulting ~1,500 nucleotide RNA is capped and polyadenylated. The 23 nucleotide single-stranded miR-155, which is harbored in exon 3, is subsequently processed from the parent RNA molecule.
The MIR155HG RNA transcript does not contain a long open reading frame (ORF), however, it does include an imperfectly base-paired stem loop that is conserved across species. This non-coding RNA (ncRNA) is now defined as a primary-miRNA (pri-miRNA). Once miR-155 pri-miRNA is transcribed, this transcript is cleaved by the nuclear microprocessor complex, of which the core components are the RNase III type endonuclease Drosha and the DiGeorge critical region 8 (DGCR8) protein, to produce a 65 nucleotide stem-loop precursor miRNA (pre-mir-155) (see Figure 2).
Following export from the nucleus by exportin-5, pre-mir-155 molecules are cleaved near the terminal loop by Dicer resulting in RNA duplexes of ~22nucleotides. Following Dicer cleavage, an Argonaute (Ago) protein binds to the short RNA duplexes, forming the core of a multi-subunit complex called the RNA-induced silencing complex (RISC). In a manner similar to siRNA duplexes, one of the two strands, the "passenger miRNA" (miR-155*), is released and degraded while the other strand, designated the "guide strand" or "mature miRNA" (miR-155), is retained within the RISC.
Recent data suggest that both arms of the pre-miRNA hairpin can give rise to mature miRNAs. Due to the increasing number of examples where two functional mature miRNAs are processed from opposite arms of the same pre-miRNA, pre-mir-155 products are now denoted with the suffix -5p (from the 5′ arm) (e.g. miR-155-5p) and -3p (from the 3′ arm) (e.g. miR-155-3p) following their name (see Figure 3).
Once miR-155-5p/-3p is assembled into the RISC, these molecules subsequently recognize their target messenger RNA (mRNA) by base pairing interactions between nucleotides 2 and 8 of miR-155-5p/-3p (the seed region) and complementary nucleotides predominantly in the 3′-untranslated region (3′-UTR) of mRNAs (see Figure 4 and 5 below). Finally, with the miR-155-5p/-3p acting as an adaptor for the RISC, complex-bound mRNAs are subjected to translational repression (i.e. inhibition of translation initiation) and/or degradation following deadenylation.
Early phylogenetic analyses demonstrated that the sequence of pre-mir-155 and miR-155-5p was conserved between human, mouse, and chicken. Recent annotated sequencing data found that 22 different organisms including, mammals, amphibians, birds, reptiles, sea squirts, and sea lampreys, express a conserved miR-155-5p.[1] Currently much less sequence data is available regarding miR-155-3p, therefore, it is not clear how conserved this miRNA is across species.[2]
Northern blot analysis found that miR-155 pri-miRNA was abundantly expressed in the human spleen and thymus and detectable in the liver, lung, and kidney. Sequence analysis of small RNA clone libraries comparing miRNA expression to all other organ systems examined established that miR-155-5p was one of five miRNAs (i.e. miR-142, miR-144, miR-150, miR-155, and miR-223) that was specific for hematopoietic cells including B-cells, T-cells, monocytes and granulocytes. Together these results suggest that miR-155-5p is expressed in a number of tissues and cell types and, therefore, may play a critical role in a wide variety of biological processes, including hematopoiesis