Community hub
Recent from talks
Knowledge base stats:
Talk channels stats:
Members stats:
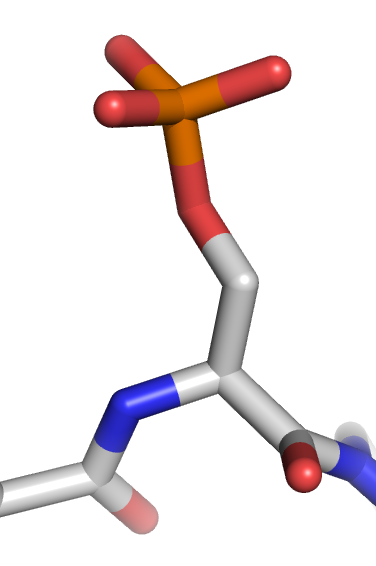
Protein phosphorylation
Protein phosphorylation is a reversible post-translational modification of proteins in which an amino acid residue is phosphorylated by a protein kinase by the addition of a covalently bound phosphate group. Phosphorylation alters the structural conformation of a protein, causing it to become activated, deactivated, or otherwise modifying its function. Approximately 13,000 human proteins have sites that are phosphorylated.
The reverse reaction of phosphorylation is called dephosphorylation, and is catalyzed by protein phosphatases. Protein kinases and phosphatases work independently and in a balance to regulate the function of proteins.
The amino acids most commonly phosphorylated are serine, threonine, tyrosine, and histidine. These phosphorylations play important and well-characterized roles in signaling pathways and metabolism. However, other amino acids can also be phosphorylated post-translationally, including arginine, lysine, aspartic acid, glutamic acid and cysteine, and these phosphorylated amino acids have been identified to be present in human cell extracts and fixed human cells using a combination of antibody-based analysis (for pHis) and mass spectrometry (for all other amino acids).
Protein phosphorylation was first reported in 1906 by Phoebus Levene at the Rockefeller Institute for Medical Research with the discovery of phosphorylated vitellin. However, it was nearly 50 years until the enzymatic phosphorylation of proteins by protein kinases was discovered.
In 1906, Phoebus Levene at the Rockefeller Institute for Medical Research identified phosphate in the protein vitellin (phosvitin) and by 1933 had detected phosphoserine in casein, with Fritz Lipmann. However, it took another 20 years before Eugene P. Kennedy described the first "enzymatic phosphorylation of proteins". The first phosphorylase enzyme was discovered by Carl and Gerty Cori in the late 1930s. Carl and Gerty Cori found two forms of glycogen phosphorylase which they named A and B but did not correctly understand the mechanism of the B form to A form conversion. The interconversion of phosphorylase b to phosphorylase a was later described by Edmond Fischer and Edwin Krebs, as well as, Wosilait and Sutherland, involving a phosphorylation/dephosphorylation mechanism. It was found that an enzyme, named phosphorylase kinase and Mg-ATP were required to phosphorylate glycogen phosphorylase by assisting in the transfer of the γ-phosphoryl group of ATP to a serine residue on phosphorylase b. Protein phosphatase 1 is able to catalyze the dephosphorylation of phosphorylated enzymes by removing the phosphate group. Earl Sutherland explained in 1950, that the activity of phosphorylase was increased and thus glycogenolysis stimulated when liver slices were incubated with adrenalin and glucagon. Phosphorylation was considered a specific control mechanism for one metabolic pathway until the 1970s, when Lester Reed discovered that mitochondrial pyruvate dehydrogenase complex was inactivated by phosphorylation. Also in the 1970s, the term multisite phosphorylation was coined in response to the discovery of proteins that are phosphorylated on two or more residues by two or more kinases. In 1975, it was shown that cAMP-dependent proteins kinases phosphorylate serine residues on specific amino acid sequence motifs. Ray Erikson discovered that v-Src was a kinase and Tony Hunter found that v-Src phosphorylated tyrosine residues on proteins in the 1970s. In the early 1980, the amino-acid sequence of the first protein kinase was determined which helped geneticists understand the functions of regulatory genes. In the late 1980s and early 1990s, the first protein tyrosine phosphatase (PTP1B) was purified and the discovery, as well as, cloning of JAK kinases was accomplished which led to many in the scientific community to name the 1990s as the decade of protein kinase cascades. Edmond Fischer and Edwin Krebs were awarded the Nobel prize in 1992 "for their discoveries concerning reversible protein phosphorylation as a biological regulatory mechanism".
Reversible phosphorylation of proteins is abundant in both prokaryotic and even more so in eukaryotic organisms. For instance, in bacteria 5–10% of all proteins are thought to be phosphorylated. By contrast, it is estimated that one third of all human proteins is phosphorylated at any point in time, with 230,000, 156,000, and 40,000 unique phosphorylation sites existing in human, mouse, and yeast, respectively. In yeast, about 120 kinases (out of ~6,000 proteins total) cause 8,814 known regulated phosphorylation events, generating about 3,600 phosphoproteins (about 60% of all yeast proteins). Hence, phosphorylation is a universal regulatory mechanism that affects a large portion of proteins. Even if a protein is not phosphorylated itself, its interactions with other proteins may be regulated by phosphorylation of these interacting proteins.
Phosphorylation introduces a charged and hydrophilic group in the side chain of amino acids, possibly changing a protein's structure by altering interactions with nearby amino acids. Some proteins such as p53 contain multiple phosphorylation sites, facilitating complex, multi-level regulation. Because of the ease with which proteins can be phosphorylated and dephosphorylated, this type of modification is a flexible mechanism for cells to respond to external signals and environmental conditions.
Kinases phosphorylate proteins and phosphatases dephosphorylate proteins. Many enzymes and receptors are switched "on" or "off" by phosphorylation and dephosphorylation. Reversible phosphorylation results in a conformational change in the structure in many enzymes and receptors, causing them to become activated or deactivated. Phosphorylation usually occurs on serine, threonine, tyrosine and histidine residues in eukaryotic proteins. Histidine phosphorylation of eukaryotic proteins appears to be much more frequent than tyrosine phosphorylation. In prokaryotic proteins phosphorylation occurs on the serine, threonine, tyrosine, histidine, arginine or lysine residues. The addition of a phosphate (PO43-) molecule to a non-polar R group of an amino acid residue can turn a hydrophobic portion of a protein into a polar and extremely hydrophilic portion of a molecule. In this way protein dynamics can induce a conformational change in the structure of the protein via long-range allostery with other hydrophobic and hydrophilic residues in the protein.
Hub AI
Protein phosphorylation AI simulator
(@Protein phosphorylation_simulator)
Protein phosphorylation
Protein phosphorylation is a reversible post-translational modification of proteins in which an amino acid residue is phosphorylated by a protein kinase by the addition of a covalently bound phosphate group. Phosphorylation alters the structural conformation of a protein, causing it to become activated, deactivated, or otherwise modifying its function. Approximately 13,000 human proteins have sites that are phosphorylated.
The reverse reaction of phosphorylation is called dephosphorylation, and is catalyzed by protein phosphatases. Protein kinases and phosphatases work independently and in a balance to regulate the function of proteins.
The amino acids most commonly phosphorylated are serine, threonine, tyrosine, and histidine. These phosphorylations play important and well-characterized roles in signaling pathways and metabolism. However, other amino acids can also be phosphorylated post-translationally, including arginine, lysine, aspartic acid, glutamic acid and cysteine, and these phosphorylated amino acids have been identified to be present in human cell extracts and fixed human cells using a combination of antibody-based analysis (for pHis) and mass spectrometry (for all other amino acids).
Protein phosphorylation was first reported in 1906 by Phoebus Levene at the Rockefeller Institute for Medical Research with the discovery of phosphorylated vitellin. However, it was nearly 50 years until the enzymatic phosphorylation of proteins by protein kinases was discovered.
In 1906, Phoebus Levene at the Rockefeller Institute for Medical Research identified phosphate in the protein vitellin (phosvitin) and by 1933 had detected phosphoserine in casein, with Fritz Lipmann. However, it took another 20 years before Eugene P. Kennedy described the first "enzymatic phosphorylation of proteins". The first phosphorylase enzyme was discovered by Carl and Gerty Cori in the late 1930s. Carl and Gerty Cori found two forms of glycogen phosphorylase which they named A and B but did not correctly understand the mechanism of the B form to A form conversion. The interconversion of phosphorylase b to phosphorylase a was later described by Edmond Fischer and Edwin Krebs, as well as, Wosilait and Sutherland, involving a phosphorylation/dephosphorylation mechanism. It was found that an enzyme, named phosphorylase kinase and Mg-ATP were required to phosphorylate glycogen phosphorylase by assisting in the transfer of the γ-phosphoryl group of ATP to a serine residue on phosphorylase b. Protein phosphatase 1 is able to catalyze the dephosphorylation of phosphorylated enzymes by removing the phosphate group. Earl Sutherland explained in 1950, that the activity of phosphorylase was increased and thus glycogenolysis stimulated when liver slices were incubated with adrenalin and glucagon. Phosphorylation was considered a specific control mechanism for one metabolic pathway until the 1970s, when Lester Reed discovered that mitochondrial pyruvate dehydrogenase complex was inactivated by phosphorylation. Also in the 1970s, the term multisite phosphorylation was coined in response to the discovery of proteins that are phosphorylated on two or more residues by two or more kinases. In 1975, it was shown that cAMP-dependent proteins kinases phosphorylate serine residues on specific amino acid sequence motifs. Ray Erikson discovered that v-Src was a kinase and Tony Hunter found that v-Src phosphorylated tyrosine residues on proteins in the 1970s. In the early 1980, the amino-acid sequence of the first protein kinase was determined which helped geneticists understand the functions of regulatory genes. In the late 1980s and early 1990s, the first protein tyrosine phosphatase (PTP1B) was purified and the discovery, as well as, cloning of JAK kinases was accomplished which led to many in the scientific community to name the 1990s as the decade of protein kinase cascades. Edmond Fischer and Edwin Krebs were awarded the Nobel prize in 1992 "for their discoveries concerning reversible protein phosphorylation as a biological regulatory mechanism".
Reversible phosphorylation of proteins is abundant in both prokaryotic and even more so in eukaryotic organisms. For instance, in bacteria 5–10% of all proteins are thought to be phosphorylated. By contrast, it is estimated that one third of all human proteins is phosphorylated at any point in time, with 230,000, 156,000, and 40,000 unique phosphorylation sites existing in human, mouse, and yeast, respectively. In yeast, about 120 kinases (out of ~6,000 proteins total) cause 8,814 known regulated phosphorylation events, generating about 3,600 phosphoproteins (about 60% of all yeast proteins). Hence, phosphorylation is a universal regulatory mechanism that affects a large portion of proteins. Even if a protein is not phosphorylated itself, its interactions with other proteins may be regulated by phosphorylation of these interacting proteins.
Phosphorylation introduces a charged and hydrophilic group in the side chain of amino acids, possibly changing a protein's structure by altering interactions with nearby amino acids. Some proteins such as p53 contain multiple phosphorylation sites, facilitating complex, multi-level regulation. Because of the ease with which proteins can be phosphorylated and dephosphorylated, this type of modification is a flexible mechanism for cells to respond to external signals and environmental conditions.
Kinases phosphorylate proteins and phosphatases dephosphorylate proteins. Many enzymes and receptors are switched "on" or "off" by phosphorylation and dephosphorylation. Reversible phosphorylation results in a conformational change in the structure in many enzymes and receptors, causing them to become activated or deactivated. Phosphorylation usually occurs on serine, threonine, tyrosine and histidine residues in eukaryotic proteins. Histidine phosphorylation of eukaryotic proteins appears to be much more frequent than tyrosine phosphorylation. In prokaryotic proteins phosphorylation occurs on the serine, threonine, tyrosine, histidine, arginine or lysine residues. The addition of a phosphate (PO43-) molecule to a non-polar R group of an amino acid residue can turn a hydrophobic portion of a protein into a polar and extremely hydrophilic portion of a molecule. In this way protein dynamics can induce a conformational change in the structure of the protein via long-range allostery with other hydrophobic and hydrophilic residues in the protein.