Recent from talks
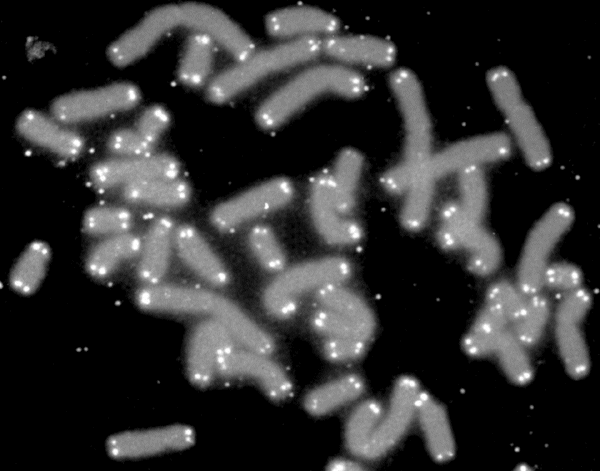
Telomere
Knowledge base stats:
Talk channels stats:
Members stats:
Telomere
A telomere (/ˈtɛləmɪər, ˈtiːlə-/; from Ancient Greek τέλος (télos) 'end' and μέρος (méros) 'part') is a region of repetitive nucleotide sequences associated with specialized proteins at the ends of linear chromosomes (see Sequences). Telomeres are a widespread genetic feature most commonly found in eukaryotes. In most or even all species possessing them, they protect the terminal regions of chromosomal DNA from progressive degradation and ensure the integrity of linear chromosomes by preventing DNA repair systems from mistaking the very ends of the DNA strand for a double-strand break.
The existence of a special structure at the ends of chromosomes was independently proposed in 1938 by Hermann Joseph Muller, studying the fruit fly Drosophila melanogaster, and in 1939 by Barbara McClintock, working with maize. Muller observed that the ends of irradiated fruit fly chromosomes did not present alterations such as deletions or inversions. He hypothesized the presence of a protective cap, which he coined "telomeres", from the Greek telos (end) and meros (part).
In the early 1970s, Soviet theorist Alexey Olovnikov first recognized that chromosomes could not completely replicate their ends; this is known as the "end replication problem". Building on this, and accommodating Leonard Hayflick's idea of limited somatic cell division, Olovnikov suggested that DNA sequences are lost every time a cell replicates until the loss reaches a critical level, at which point cell division ends. According to his theory of marginotomy, DNA sequences at the ends of telomeres are represented by tandem repeats, which create a buffer that determines the number of divisions that a certain cell clone can undergo. Furthermore, it was predicted that a specialized DNA polymerase (originally called a tandem-DNA-polymerase) could extend telomeres in immortal tissues such as germ line, cancer cells and stem cells. It also followed from this hypothesis that organisms with circular genome, such as bacteria, do not have the end replication problem and therefore do not age.
Olovnikov suggested that in germline cells, cells of vegetatively propagated organisms, and immortal cell populations such as most cancer cell lines, an enzyme might be activated to prevent the shortening of DNA termini with each cell division.
In 1975–1977, Elizabeth Blackburn, working as a postdoctoral fellow at Yale University with Joseph G. Gall, discovered the unusual nature of telomeres, with their simple repeated DNA sequences composing chromosome ends. Blackburn, Carol Greider, and Jack Szostak were awarded the 2009 Nobel Prize in Physiology or Medicine for the discovery of how chromosomes are protected by telomeres and the enzyme telomerase.
During DNA replication, DNA polymerase cannot replicate the sequences present at the 3' ends of the parent strands. This is a consequence of its unidirectional mode of DNA synthesis: it can only attach new nucleotides to an existing 3'-end (that is, synthesis progresses 5'-3') and thus it requires a primer to initiate replication. On the leading strand (oriented 5'-3' within the replication fork), DNA-polymerase continuously replicates from the point of initiation all the way to the strand's end with the primer (made of RNA) then being excised and substituted by DNA. The lagging strand, however, is oriented 3'-5' with respect to the replication fork so continuous replication by DNA-polymerase is impossible, which necessitates discontinuous replication involving the repeated synthesis of primers further 5' of the site of initiation (see lagging strand replication). The last primer to be involved in lagging-strand replication sits near the 3'-end of the template (corresponding to the potential 5'-end of the lagging-strand). Originally it was believed that the last primer would sit at the very end of the template, thus, once removed, the DNA-polymerase that substitutes primers with DNA (DNA-Pol δ in eukaryotes) would be unable to synthesize the "replacement DNA" from the 5'-end of the lagging strand so that the template nucleotides previously paired to the last primer would not be replicated. It has since been questioned whether the last lagging strand primer is placed exactly at the 3'-end of the template and it was demonstrated that it is rather synthesized at a distance of about 70–100 nucleotides which is consistent with the finding that DNA in cultured human cell is shortened by 50–100 base pairs per cell division.
If coding sequences are degraded in this process, potentially vital genetic information would be lost. Telomeres are non-coding, repetitive sequences located at the termini of linear chromosomes to act as buffers for those coding sequences further behind. They "cap" the end-sequences and are progressively degraded in the process of DNA replication.
The "end replication problem" is exclusive to linear chromosomes as circular chromosomes do not have ends lying without reach of DNA-polymerases. Most prokaryotes, relying on circular chromosomes, accordingly do not possess telomeres. A small fraction of bacterial chromosomes (such as those in Streptomyces, Agrobacterium, and Borrelia), however, are linear and possess telomeres, which are very different from those of the eukaryotic chromosomes in structure and function. The known structures of bacterial telomeres take the form of proteins bound to the ends of linear chromosomes, or hairpin loops of single-stranded DNA at the ends of the linear chromosomes.
Hub AI
Telomere AI simulator
(@Telomere_simulator)
Telomere
A telomere (/ˈtɛləmɪər, ˈtiːlə-/; from Ancient Greek τέλος (télos) 'end' and μέρος (méros) 'part') is a region of repetitive nucleotide sequences associated with specialized proteins at the ends of linear chromosomes (see Sequences). Telomeres are a widespread genetic feature most commonly found in eukaryotes. In most or even all species possessing them, they protect the terminal regions of chromosomal DNA from progressive degradation and ensure the integrity of linear chromosomes by preventing DNA repair systems from mistaking the very ends of the DNA strand for a double-strand break.
The existence of a special structure at the ends of chromosomes was independently proposed in 1938 by Hermann Joseph Muller, studying the fruit fly Drosophila melanogaster, and in 1939 by Barbara McClintock, working with maize. Muller observed that the ends of irradiated fruit fly chromosomes did not present alterations such as deletions or inversions. He hypothesized the presence of a protective cap, which he coined "telomeres", from the Greek telos (end) and meros (part).
In the early 1970s, Soviet theorist Alexey Olovnikov first recognized that chromosomes could not completely replicate their ends; this is known as the "end replication problem". Building on this, and accommodating Leonard Hayflick's idea of limited somatic cell division, Olovnikov suggested that DNA sequences are lost every time a cell replicates until the loss reaches a critical level, at which point cell division ends. According to his theory of marginotomy, DNA sequences at the ends of telomeres are represented by tandem repeats, which create a buffer that determines the number of divisions that a certain cell clone can undergo. Furthermore, it was predicted that a specialized DNA polymerase (originally called a tandem-DNA-polymerase) could extend telomeres in immortal tissues such as germ line, cancer cells and stem cells. It also followed from this hypothesis that organisms with circular genome, such as bacteria, do not have the end replication problem and therefore do not age.
Olovnikov suggested that in germline cells, cells of vegetatively propagated organisms, and immortal cell populations such as most cancer cell lines, an enzyme might be activated to prevent the shortening of DNA termini with each cell division.
In 1975–1977, Elizabeth Blackburn, working as a postdoctoral fellow at Yale University with Joseph G. Gall, discovered the unusual nature of telomeres, with their simple repeated DNA sequences composing chromosome ends. Blackburn, Carol Greider, and Jack Szostak were awarded the 2009 Nobel Prize in Physiology or Medicine for the discovery of how chromosomes are protected by telomeres and the enzyme telomerase.
During DNA replication, DNA polymerase cannot replicate the sequences present at the 3' ends of the parent strands. This is a consequence of its unidirectional mode of DNA synthesis: it can only attach new nucleotides to an existing 3'-end (that is, synthesis progresses 5'-3') and thus it requires a primer to initiate replication. On the leading strand (oriented 5'-3' within the replication fork), DNA-polymerase continuously replicates from the point of initiation all the way to the strand's end with the primer (made of RNA) then being excised and substituted by DNA. The lagging strand, however, is oriented 3'-5' with respect to the replication fork so continuous replication by DNA-polymerase is impossible, which necessitates discontinuous replication involving the repeated synthesis of primers further 5' of the site of initiation (see lagging strand replication). The last primer to be involved in lagging-strand replication sits near the 3'-end of the template (corresponding to the potential 5'-end of the lagging-strand). Originally it was believed that the last primer would sit at the very end of the template, thus, once removed, the DNA-polymerase that substitutes primers with DNA (DNA-Pol δ in eukaryotes) would be unable to synthesize the "replacement DNA" from the 5'-end of the lagging strand so that the template nucleotides previously paired to the last primer would not be replicated. It has since been questioned whether the last lagging strand primer is placed exactly at the 3'-end of the template and it was demonstrated that it is rather synthesized at a distance of about 70–100 nucleotides which is consistent with the finding that DNA in cultured human cell is shortened by 50–100 base pairs per cell division.
If coding sequences are degraded in this process, potentially vital genetic information would be lost. Telomeres are non-coding, repetitive sequences located at the termini of linear chromosomes to act as buffers for those coding sequences further behind. They "cap" the end-sequences and are progressively degraded in the process of DNA replication.
The "end replication problem" is exclusive to linear chromosomes as circular chromosomes do not have ends lying without reach of DNA-polymerases. Most prokaryotes, relying on circular chromosomes, accordingly do not possess telomeres. A small fraction of bacterial chromosomes (such as those in Streptomyces, Agrobacterium, and Borrelia), however, are linear and possess telomeres, which are very different from those of the eukaryotic chromosomes in structure and function. The known structures of bacterial telomeres take the form of proteins bound to the ends of linear chromosomes, or hairpin loops of single-stranded DNA at the ends of the linear chromosomes.
Recent media