Community hub
Doublecortin
View on Wikipedia| DCX | |||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 | |||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||
| Identifiers | |||||||||||||||||||||||||||||||
| Aliases | DCX, DBCN, DC, LISX, SCLH, XLIS, doublecortin | ||||||||||||||||||||||||||||||
| External IDs | OMIM: 300121; MGI: 1277171; HomoloGene: 7683; GeneCards: DCX; OMA:DCX - orthologs | ||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||
| Wikidata | |||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||






Neuronal migration protein doublecortin, also known as doublin or lissencephalin-X is a protein that in humans is encoded by the DCX gene.[5]
Function
[edit]
Doublecortin (DCX) is a microtubule-associated protein expressed by neuronal precursor cells and immature neurons in embryonic and adult cortical structures. Neuronal precursor cells begin to express DCX while actively dividing, and their neuronal daughter cells continue to express DCX for 2–3 weeks as the cells mature into neurons. Downregulation of DCX begins after 2 weeks, and occurs at the same time that these cells begin to express NeuN, a neuronal marker.[7]
Due to the nearly exclusive expression of DCX in developing neurons, this protein has been used increasingly as a marker for neurogenesis. Indeed, levels of DCX expression increase in response to exercise,[8] and that increase occurs in parallel with increased BrdU labeling, which is currently a "gold standard" in measuring neurogenesis.
Doublecortin was found to bind to the microtubule cytoskeleton. In vivo and in vitro assays show that Doublecortin stabilizes microtubules and causes bundling.[9] Doublecortin is a basic protein with an iso-electric point of 10 typical of microtubule-binding proteins.
Knock out mouse
[edit]
In mice where the Doublecortin gene has been knocked out, cortical layers are still correctly formed. However, the hippocampi of these mice show disorganisation in the CA3 region. The normally single layer of pyramidal cells in mutants is seen as a double layer. These mice also have different behavior than their wild type littermates and are epileptic.[10]
Structure
[edit]| Doublecortin | |||||||||
|---|---|---|---|---|---|---|---|---|---|
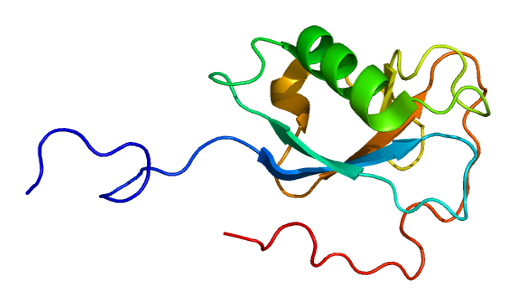
 solution structure of the N-terminal dcx domain of human doublecortin-like kinase | |||||||||
| Identifiers | |||||||||
| Symbol | DCX | ||||||||
| Pfam | PF03607 | ||||||||
| InterPro | IPR003533 | ||||||||
| SCOP2 | 1mfw / SCOPe / SUPFAM | ||||||||
| CDD | cd01617 | ||||||||
| |||||||||
The detailed sequence analysis of Doublecortin and Doublecortin-like proteins allowed the identification of a tandem repeat of evolutionarily conserved Doublecortin (DC) domains. These domains are found in the N terminus of proteins and consists of tandemly repeated copies of an around 80 amino acids region. It has been suggested that the first DC domain of Doublecortin binds tubulin and enhances microtubule polymerisation.[11]
Doublecortin has been shown to influence the structure of microtubules. Microtubule nucleated in vitro in the presence of Doublecortin have almost exclusively 13 protofilaments, whereas microtubule nucleated without Doublecortin are present in a range of different sizes.
Interactions
[edit]Clinical significance
[edit]Doublecortin is mutated in X-linked lissencephaly and the double cortex syndrome, and the clinical manifestations are sex-linked. In males, X-linked lissencephaly produces a smooth brain due to lack of migration of immature neurons, which normally promote folding of the brain surface. Double cortex syndrome is characterized by abnormal migration of neural tissue during development which results in two bands of misplaced neurons within the subcortical white, generating two cortices, giving the name to the syndrome; this finding generally occurs in females.[13] The mutation was discovered by Joseph Gleeson and Christopher A. Walsh in Boston.[14][15] At least 49 disease-causing mutations in this gene have been discovered.[16]
See also
[edit]References
[edit]- ^ a b c GRCh38: Ensembl release 89: ENSG00000077279 – Ensembl, May 2017
- ^ a b c GRCm38: Ensembl release 89: ENSMUSG00000031285 – Ensembl, May 2017
- ^ "Human PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- ^ "Mouse PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- ^ EntrezGene 1641
- ^ Oomen CA, Girardi CE, Cahyadi R, Verbeek EC, Krugers H, Joëls M, Lucassen PJ (2009). "Opposite effects of early maternal deprivation on neurogenesis in male versus female rats". PLOS ONE. 4 (1) e3675. Bibcode:2009PLoSO...4.3675O. doi:10.1371/journal.pone.0003675. PMC 2629844. PMID 19180242.
- ^ Brown JP, Couillard-Després S, Cooper-Kuhn CM, Winkler J, Aigner L, Kuhn HG (December 2003). "Transient expression of doublecortin during adult neurogenesis". J. Comp. Neurol. 467 (1): 1–10. doi:10.1002/cne.10874. PMID 14574675. S2CID 25315374.
- ^ Couillard-Despres S, Winner B, Schaubeck S, Aigner R, Vroemen M, Weidner N, Bogdahn U, Winkler J, Kuhn HG, Aigner L (January 2005). "Doublecortin expression levels in adult brain reflect neurogenesis". Eur. J. Neurosci. 21 (1): 1–14. doi:10.1111/j.1460-9568.2004.03813.x. PMID 15654838. S2CID 45893767.
- ^ Horesh D, Sapir T, Francis F, Wolf SG, Caspi M, Elbaum M, Chelly J, Reiner O (September 1999). "Doublecortin, a stabilizer of microtubules". Hum. Mol. Genet. 8 (9): 1599–610. doi:10.1093/hmg/8.9.1599. PMID 10441322.
- ^ Nosten-Bertrand M, Kappeler C, Dinocourt C, Denis C, Germain J, Phan Dinh Tuy F, Verstraeten S, Alvarez C, Métin C, Chelly J, Giros B, Miles R, Depaulis A, Francis F (2008-06-25). "Epilepsy in Dcx knockout mice associated with discrete lamination defects and enhanced excitability in the hippocampus". PLOS ONE. 3 (6) e2473. Bibcode:2008PLoSO...3.2473N. doi:10.1371/journal.pone.0002473. PMC 2429962. PMID 18575605.
- ^ Sapir T, Horesh D, Caspi M, Atlas R, Burgess HA, Wolf SG, Francis F, Chelly J, Elbaum M, Pietrokovski S, Reiner O (March 2000). "Doublecortin mutations cluster in evolutionarily conserved functional domains". Hum. Mol. Genet. 9 (5): 703–12. doi:10.1093/hmg/9.5.703. PMID 10749977.
- ^ Caspi M, Atlas R, Kantor A, Sapir T, Reiner O (September 2000). "Interaction between LIS1 and doublecortin, two lissencephaly gene products". Hum. Mol. Genet. 9 (15): 2205–13. doi:10.1093/oxfordjournals.hmg.a018911. PMID 11001923.
- ^ Online Mendelian Inheritance in Man (OMIM): Doublecortin - 300121
- ^ Gleeson JG, Allen KM, Fox JW, Lamperti ED, Berkovic S, Scheffer I, Cooper EC, Dobyns WB, Minnerath SR, Ross ME, Walsh CA (January 1998). "Doublecortin, a brain-specific gene mutated in human X-linked lissencephaly and double cortex syndrome, encodes a putative signaling protein". Cell. 92 (1): 63–72. doi:10.1016/S0092-8674(00)80899-5. PMID 9489700.
- ^ Lowenstein DH (2011). "Seizures and Epilepsy". In Loscalzo J, Longo DL, Fauci AS, Kasper DL, Hauser SL (eds.). Harrison's Principles of Internal Medicine (18th ed.). McGraw-Hill Professional. pp. 3251–3269. ISBN 978-0-07-174889-6.
- ^ Šimčíková D, Heneberg P (December 2019). "Refinement of evolutionary medicine predictions based on clinical evidence for the manifestations of Mendelian diseases". Scientific Reports. 9 (1) 18577. Bibcode:2019NatSR...918577S. doi:10.1038/s41598-019-54976-4. PMC 6901466. PMID 31819097.
Further reading
[edit]- des Portes V, Pinard JM, Smadja D, et al. (1997). "Dominant X linked subcortical laminar heterotopia and lissencephaly syndrome (XSCLH/LIS): evidence for the occurrence of mutation in males and mapping of a potential locus in Xq22". J. Med. Genet. 34 (3): 177–83. doi:10.1136/jmg.34.3.177. PMC 1050888. PMID 9132485.
- des Portes V, Pinard JM, Billuart P, et al. (1998). "A novel CNS gene required for neuronal migration and involved in X-linked subcortical laminar heterotopia and lissencephaly syndrome". Cell. 92 (1): 51–61. doi:10.1016/S0092-8674(00)80898-3. PMID 9489699.
- Gleeson JG, Allen KM, Fox JW, et al. (1998). "Doublecortin, a brain-specific gene mutated in human X-linked lissencephaly and double cortex syndrome, encodes a putative signaling protein". Cell. 92 (1): 63–72. doi:10.1016/S0092-8674(00)80899-5. PMID 9489700.
- des Portes V, Francis F, Pinard JM, et al. (1999). "doublecortin is the major gene causing X-linked subcortical laminar heterotopia (SCLH)". Hum. Mol. Genet. 7 (7): 1063–70. doi:10.1093/hmg/7.7.1063. PMID 9618162.
- Sossey-Alaoui K, Hartung AJ, Guerrini R, et al. (1998). "Human doublecortin (DCX) and the homologous gene in mouse encode a putative Ca2+-dependent signaling protein which is mutated in human X-linked neuronal migration defects". Hum. Mol. Genet. 7 (8): 1327–32. doi:10.1093/hmg/7.8.1327. PMID 9668176.
- Pilz DT, Matsumoto N, Minnerath S, et al. (1999). "LIS1 and XLIS (DCX) mutations cause most classical lissencephaly, but different patterns of malformation". Hum. Mol. Genet. 7 (13): 2029–37. doi:10.1093/hmg/7.13.2029. PMID 9817918.
- Gleeson JG, Minnerath SR, Fox JW, et al. (1999). "Characterization of mutations in the gene doublecortin in patients with double cortex syndrome". Ann. Neurol. 45 (2): 146–53. doi:10.1002/1531-8249(199902)45:2<146::AID-ANA3>3.0.CO;2-N. PMID 9989615. S2CID 37046760.
- Kato M, Kimura T, Lin C, et al. (1999). "A novel mutation of the doublecortin gene in Japanese patients with X-linked lissencephaly and subcortical band heterotopia". Hum. Genet. 104 (4): 341–4. doi:10.1007/s004390050963. PMID 10369164. S2CID 875280.
- Gleeson JG, Lin PT, Flanagan LA, Walsh CA (1999). "Doublecortin is a microtubule-associated protein and is expressed widely by migrating neurons". Neuron. 23 (2): 257–71. doi:10.1016/S0896-6273(00)80778-3. PMID 10399933.
- Pilz DT, Kuc J, Matsumoto N, et al. (2000). "Subcortical band heterotopia in rare affected males can be caused by missense mutations in DCX (XLIS) or LIS1". Hum. Mol. Genet. 8 (9): 1757–60. doi:10.1093/hmg/8.9.1757. PMID 10441340.
- Sakamoto M, Ono J, Okada S, et al. (2000). "Genetic alteration of the DCX gene in Japanese patients with subcortical laminar heterotopia or isolated lissencephaly sequence". J. Hum. Genet. 45 (3): 167–70. doi:10.1007/s100380050204. PMID 10807542.
- Caspi M, Atlas R, Kantor A, et al. (2001). "Interaction between LIS1 and doublecortin, two lissencephaly gene products". Hum. Mol. Genet. 9 (15): 2205–13. doi:10.1093/oxfordjournals.hmg.a018911. PMID 11001923.
- Matsumoto N, Leventer RJ, Kuc JA, et al. (2001). "Mutation analysis of the DCX gene and genotype/phenotype correlation in subcortical band heterotopia". Eur. J. Hum. Genet. 9 (1): 5–12. doi:10.1038/sj.ejhg.5200548. PMID 11175293.
- Demelas L, Serra G, Conti M, et al. (2001). "Incomplete penetrance with normal MRI in a woman with germline mutation of the DCX gene". Neurology. 57 (2): 327–30. doi:10.1212/wnl.57.2.327. PMID 11468322. S2CID 6735445.
- Friocourt G, Chafey P, Billuart P, et al. (2001). "Doublecortin interacts with mu subunits of clathrin adaptor complexes in the developing nervous system". Mol. Cell. Neurosci. 18 (3): 307–19. doi:10.1006/mcne.2001.1022. PMID 11591131. S2CID 54264008.
- Kato M, Kanai M, Soma O, et al. (2001). "Mutation of the doublecortin gene in male patients with double cortex syndrome: somatic mosaicism detected by hair root analysis". Ann. Neurol. 50 (4): 547–51. doi:10.1002/ana.1231. PMID 11601509. S2CID 18259219.
- des Portes V, Abaoub L, Joannard A, et al. (2002). "So-called 'cryptogenic' partial seizures resulting from a subtle cortical dysgenesis due to a doublecortin gene mutation". Seizure: The Journal of the British Epilepsy Association. 11 (4): 273–7. doi:10.1053/seiz.2001.0607. PMID 12027577.
- Kizhatil K, Wu YX, Sen A, Bennett V (2002). "A new activity of doublecortin in recognition of the phospho-FIGQY tyrosine in the cytoplasmic domain of neurofascin". J. Neurosci. 22 (18): 7948–58. doi:10.1523/JNEUROSCI.22-18-07948.2002. PMC 6758080. PMID 12223548.
- D'Agostino MD, Bernasconi A, Das S, et al. (2002). "Subcortical band heterotopia (SBH) in males: clinical, imaging and genetic findings in comparison with females". Brain. 125 (Pt 11): 2507–22. doi:10.1093/brain/awf248. hdl:10923/21721. PMID 12390976.
- Meyer G, Perez-Garcia CG, Gleeson JG (2003). "Selective expression of doublecortin and LIS1 in developing human cortex suggests unique modes of neuronal movement". Cereb. Cortex. 12 (12): 1225–36. doi:10.1093/cercor/12.12.1225. PMID 12427674.
External links
[edit] Media related to Doublecortin at Wikimedia Commons
Media related to Doublecortin at Wikimedia Commons- GeneReviews/NCBI/NIH/UW entry on DCX-Related Disorders
- OMIM entries on DCX-Related Disorders
- doublecortin+protein at the U.S. National Library of Medicine Medical Subject Headings (MeSH)
- Overview of all the structural information available in the PDB for UniProt: O43602 (Neuronal migration protein doublecortin) at the PDBe-KB.
PDB gallery | |
|---|---|
|

