
Community hub
Recent from talks
Contribute something to knowledge base
Content stats: 0 posts, 0 articles, 1 media, 0 notes
Members stats: 0 subscribers, 0 contributors, 0 moderators, 0 supporters
Subscribers
Supporters
Contributors
Moderators
Hub AI
Nuclear lamina AI simulator
(@Nuclear lamina_simulator)
Hub AI
Nuclear lamina AI simulator
(@Nuclear lamina_simulator)
Nuclear lamina
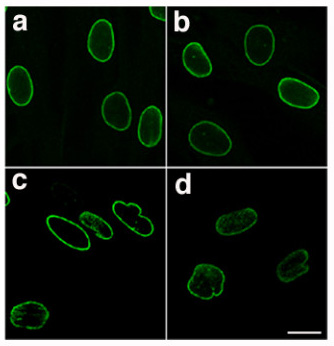
The nuclear lamina is a dense (~30 to 100 nm thick) fibrillar network inside the nucleus of eukaryote cells. It is composed of intermediate filaments and membrane associated proteins. Besides providing mechanical support, the nuclear lamina regulates important cellular events such as DNA replication and cell division. Additionally, it participates in chromatin organization and it anchors the nuclear pore complexes embedded in the nuclear envelope.
The nuclear lamina is associated with the inner face of the inner nuclear membrane of the nuclear envelope, whereas the outer face of the outer nuclear membrane is continuous with the endoplasmic reticulum. The nuclear lamina is similar in structure to the nuclear matrix, that extends throughout the nucleoplasm.
The nuclear lamina consists of two components, lamins and nuclear lamin-associated membrane proteins. The lamins are type V intermediate filaments which can be categorized as either A-type (lamin A, C) or B-type (lamin B1, B2) according to homology of their DNA sequences, biochemical properties and cellular localization during the cell cycle. Type V intermediate filaments differ from cytoplasmic intermediate filaments in the way that they have an extended rod domain (42 amino acid longer), that they all carry a nuclear localization signal (NLS) at their C-terminus and that they display typical tertiary structures. Lamin polypeptides have an almost complete α-helical conformation with multiple α-helical domains separated by non-α-helical linkers that are highly conserved in length and amino acid sequence. Both the C-terminus and the N-terminus are non α-helical, with the C-terminus displaying a globular structure with immunoglobulin type folded motif. Their molecular weight ranges from 60 to 80 kilodaltons (kDa). In the amino acid sequence of a nuclear lamin, there are also two phosphoacceptor sites present, flanking the central rod domain. A phosphorylation event at the onset of mitosis leads to a conformational change which causes the disassembly of the nuclear lamina. (discussed later in the article)
In the vertebrate genome, lamins are encoded by three genes. By alternative splicing, at least seven different polypeptides (splice variants) are obtained, some of which are specific for germ cells and play an important role in the chromatin reorganisation during meiosis. Not all organisms have the same number of lamin encoding genes; Drosophila melanogaster for example has only 2 genes, whereas Caenorhabditis elegans has only one.
The presence of lamin polypeptides is a property of all animals.
The nuclear lamin-associated membrane proteins are either integral or peripheral membrane proteins. The most important are lamina associated polypeptides 1 and 2 (LAP1, LAP2), emerin, lamin B-receptor (LBR), otefin and MAN1. Due to their positioning within or their association with the inner membrane, they mediate the attachment of the nuclear lamina to the nuclear envelope.
The nuclear lamina is assembled by interactions of two lamin polypeptides in which the α-helical regions are wound around each other to form a two stranded α-helical coiled-coil structure, followed by a head-to-tail association of the multiple dimers. The linearly elongated polymer is extended laterally by a side-by-side association of polymers, resulting in a 2D structure underlying the nuclear envelope. Next to providing mechanical support to the nucleus, the nuclear lamina plays an essential role in chromatin organization, cell cycle regulation, DNA replication, DNA repair, cell differentiation and apoptosis.
The non-random organization of the genome strongly suggests that the nuclear lamina plays a role in chromatin organization. It has been shown that lamin polypeptides have an affinity for binding chromatin through their α-helical (rod like) domains at specific DNA sequences called matrix attachment regions (MAR). A MAR has a length of approximately 300–1000 bp and has a high A/T content. Lamin A and B can also bind core histones through a sequence element in their tail domain.
Nuclear lamina
The nuclear lamina is a dense (~30 to 100 nm thick) fibrillar network inside the nucleus of eukaryote cells. It is composed of intermediate filaments and membrane associated proteins. Besides providing mechanical support, the nuclear lamina regulates important cellular events such as DNA replication and cell division. Additionally, it participates in chromatin organization and it anchors the nuclear pore complexes embedded in the nuclear envelope.
The nuclear lamina is associated with the inner face of the inner nuclear membrane of the nuclear envelope, whereas the outer face of the outer nuclear membrane is continuous with the endoplasmic reticulum. The nuclear lamina is similar in structure to the nuclear matrix, that extends throughout the nucleoplasm.
The nuclear lamina consists of two components, lamins and nuclear lamin-associated membrane proteins. The lamins are type V intermediate filaments which can be categorized as either A-type (lamin A, C) or B-type (lamin B1, B2) according to homology of their DNA sequences, biochemical properties and cellular localization during the cell cycle. Type V intermediate filaments differ from cytoplasmic intermediate filaments in the way that they have an extended rod domain (42 amino acid longer), that they all carry a nuclear localization signal (NLS) at their C-terminus and that they display typical tertiary structures. Lamin polypeptides have an almost complete α-helical conformation with multiple α-helical domains separated by non-α-helical linkers that are highly conserved in length and amino acid sequence. Both the C-terminus and the N-terminus are non α-helical, with the C-terminus displaying a globular structure with immunoglobulin type folded motif. Their molecular weight ranges from 60 to 80 kilodaltons (kDa). In the amino acid sequence of a nuclear lamin, there are also two phosphoacceptor sites present, flanking the central rod domain. A phosphorylation event at the onset of mitosis leads to a conformational change which causes the disassembly of the nuclear lamina. (discussed later in the article)
In the vertebrate genome, lamins are encoded by three genes. By alternative splicing, at least seven different polypeptides (splice variants) are obtained, some of which are specific for germ cells and play an important role in the chromatin reorganisation during meiosis. Not all organisms have the same number of lamin encoding genes; Drosophila melanogaster for example has only 2 genes, whereas Caenorhabditis elegans has only one.
The presence of lamin polypeptides is a property of all animals.
The nuclear lamin-associated membrane proteins are either integral or peripheral membrane proteins. The most important are lamina associated polypeptides 1 and 2 (LAP1, LAP2), emerin, lamin B-receptor (LBR), otefin and MAN1. Due to their positioning within or their association with the inner membrane, they mediate the attachment of the nuclear lamina to the nuclear envelope.
The nuclear lamina is assembled by interactions of two lamin polypeptides in which the α-helical regions are wound around each other to form a two stranded α-helical coiled-coil structure, followed by a head-to-tail association of the multiple dimers. The linearly elongated polymer is extended laterally by a side-by-side association of polymers, resulting in a 2D structure underlying the nuclear envelope. Next to providing mechanical support to the nucleus, the nuclear lamina plays an essential role in chromatin organization, cell cycle regulation, DNA replication, DNA repair, cell differentiation and apoptosis.
The non-random organization of the genome strongly suggests that the nuclear lamina plays a role in chromatin organization. It has been shown that lamin polypeptides have an affinity for binding chromatin through their α-helical (rod like) domains at specific DNA sequences called matrix attachment regions (MAR). A MAR has a length of approximately 300–1000 bp and has a high A/T content. Lamin A and B can also bind core histones through a sequence element in their tail domain.
Recent media
Recent media