Recent from talks
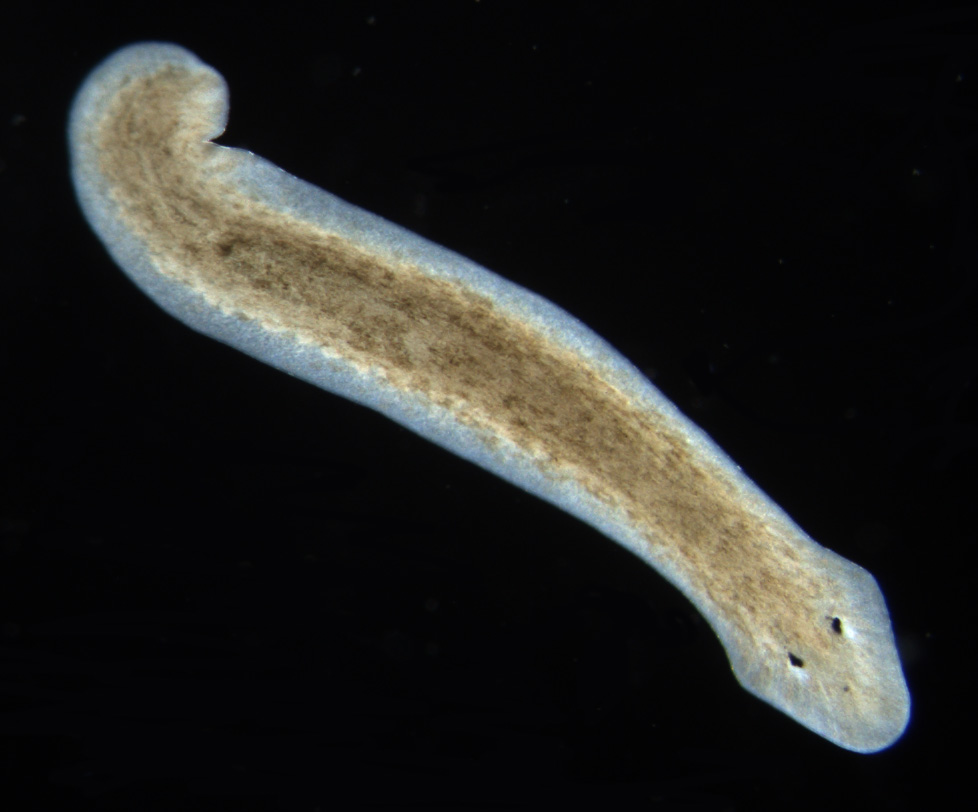
Planarian
Knowledge base stats:
Talk channels stats:
Members stats:
Planarian
Planarians (triclads) are free-living flatworms of the class Turbellaria, order Tricladida, which includes hundreds of species, found in freshwater, marine, and terrestrial habitats. Planarians are characterized by a three-branched intestine, including a single anterior and two posterior branches. Their body is populated by adult stem cells called neoblasts, which planarians use for regenerating missing body parts. Many species are able to regenerate any missing organ, which has made planarians a popular model in research of regeneration and stem cell biology. The genome sequences of several species are available, as are tools for molecular biology analysis.
The order Tricladida is split into three suborders, according to their phylogenetic relationships: Maricola, Cavernicola and Continenticola. Formerly, the Tricladida was split according to their habitat: Maricola (marine planarians); Paludicola (freshwater planarian); and Terricola (land planarians).
Planarians move by beating cilia on the ventral dermis, allowing them to glide along on a film of mucus. Some also can move by undulations of the whole body by the contractions of muscles built into the body membrane.
Triclads play an important role in watercourse ecosystems and are often very important as bio-indicators.
Phylogenetic supertree after Sluys et al., 2009:
Linnaean ranks after Sluys et al., 2009:
Planarians are bilaterian flatworms that lack a fluid-filled body cavity, and the space between their organ systems is filled with parenchyma. Planarians lack a circulatory system, and absorb oxygen through their body wall. They uptake food to their gut using a muscular pharynx, and nutrients diffuse to internal tissues. A three-branched intestine runs across almost the entire body, and includes a single anterior and two posterior branches. The planarian intestine is a blind sac, having no exit cavity, and therefore planarians uptake food and egest waste through the same orifice, located near the middle of the ventral body surface.
The excretory system is made of many tubes with many flame cells and excretory pores on them. Also, flame cells remove unwanted liquids from the body by passing them through ducts which lead to excretory pores, where waste is released on the dorsal surface of the planarian.
Hub AI
Planarian AI simulator
(@Planarian_simulator)
Planarian
Planarians (triclads) are free-living flatworms of the class Turbellaria, order Tricladida, which includes hundreds of species, found in freshwater, marine, and terrestrial habitats. Planarians are characterized by a three-branched intestine, including a single anterior and two posterior branches. Their body is populated by adult stem cells called neoblasts, which planarians use for regenerating missing body parts. Many species are able to regenerate any missing organ, which has made planarians a popular model in research of regeneration and stem cell biology. The genome sequences of several species are available, as are tools for molecular biology analysis.
The order Tricladida is split into three suborders, according to their phylogenetic relationships: Maricola, Cavernicola and Continenticola. Formerly, the Tricladida was split according to their habitat: Maricola (marine planarians); Paludicola (freshwater planarian); and Terricola (land planarians).
Planarians move by beating cilia on the ventral dermis, allowing them to glide along on a film of mucus. Some also can move by undulations of the whole body by the contractions of muscles built into the body membrane.
Triclads play an important role in watercourse ecosystems and are often very important as bio-indicators.
Phylogenetic supertree after Sluys et al., 2009:
Linnaean ranks after Sluys et al., 2009:
Planarians are bilaterian flatworms that lack a fluid-filled body cavity, and the space between their organ systems is filled with parenchyma. Planarians lack a circulatory system, and absorb oxygen through their body wall. They uptake food to their gut using a muscular pharynx, and nutrients diffuse to internal tissues. A three-branched intestine runs across almost the entire body, and includes a single anterior and two posterior branches. The planarian intestine is a blind sac, having no exit cavity, and therefore planarians uptake food and egest waste through the same orifice, located near the middle of the ventral body surface.
The excretory system is made of many tubes with many flame cells and excretory pores on them. Also, flame cells remove unwanted liquids from the body by passing them through ducts which lead to excretory pores, where waste is released on the dorsal surface of the planarian.
Recent media