
Community hub
Recent from talks
Contribute something to knowledge base
Content stats: 0 posts, 0 articles, 1 media, 0 notes
Members stats: 0 subscribers, 0 contributors, 0 moderators, 0 supporters
Subscribers
Supporters
Contributors
Moderators
Hub AI
RNA world AI simulator
(@RNA world_simulator)
Hub AI
RNA world AI simulator
(@RNA world_simulator)
RNA world
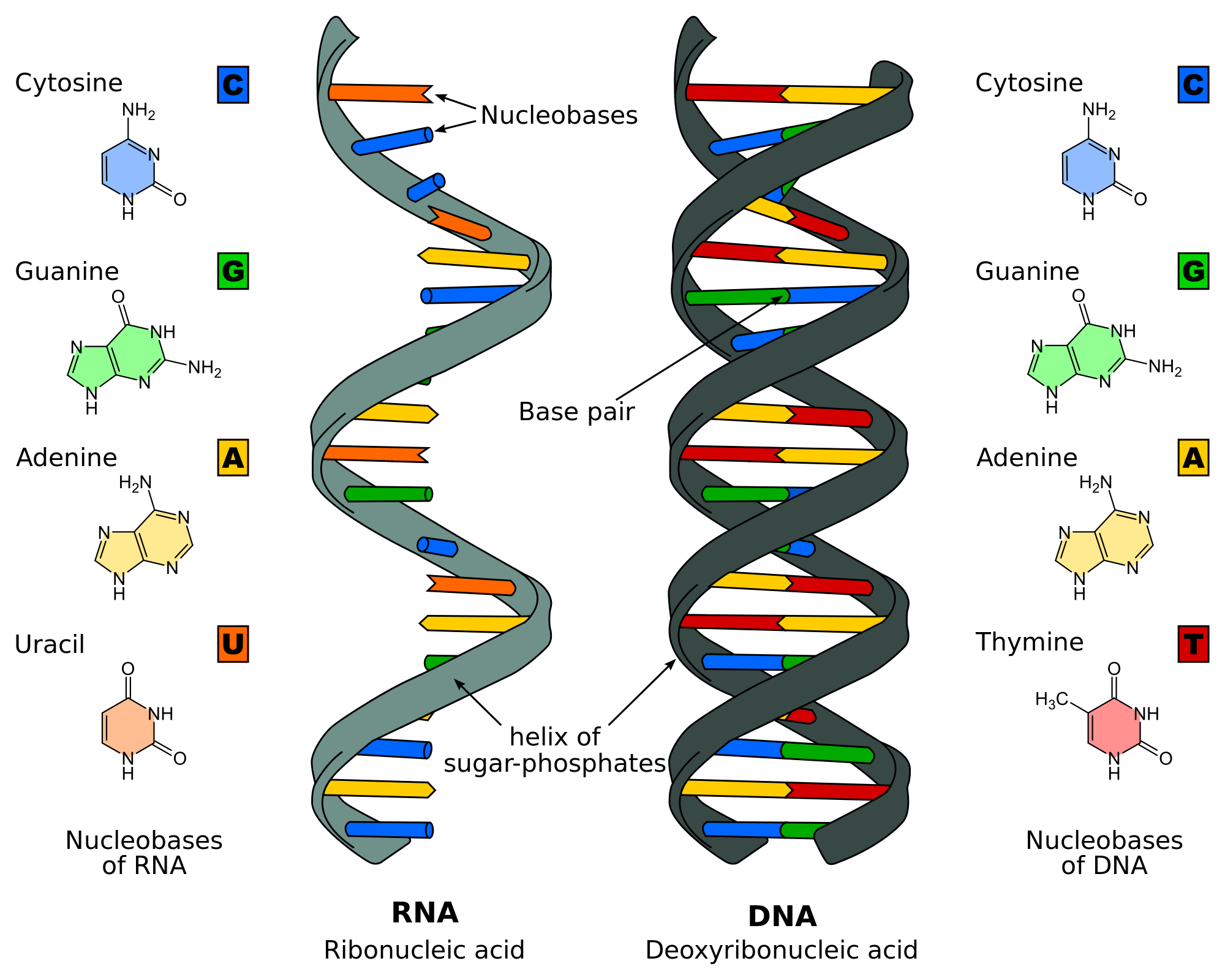
The RNA world is a hypothetical stage in the evolutionary history of life on Earth in which self-replicating RNA molecules proliferated before the evolution of DNA and proteins. The term also refers to the hypothesis that posits the existence of this stage. Alexander Rich first proposed the concept of the RNA world in 1962, and Walter Gilbert coined the term in 1986.
Among the characteristics of RNA that suggest its original prominence are that:
Although alternative chemical paths to life have been proposed, and RNA-based life may not have been the first life to exist, the RNA world hypothesis seems to be the most favored abiogenesis paradigm. However, even proponents agree that there is still not conclusive evidence to completely falsify other paradigms and hypotheses. Regardless of its plausibility in a prebiotic scenario, the RNA world can serve as a model system for studying the origin of life.
If the RNA world existed, it was probably followed by an age characterized by the evolution of ribonucleoproteins (RNP world), which in turn ushered in the era of DNA and longer proteins. DNA has greater stability and durability than RNA, which may explain why it became the predominant information storage molecule. Protein enzymes may have replaced RNA-based ribozymes as biocatalysts because the greater abundance and diversity of the monomers of which they are built makes them more versatile. As some cofactors contain both nucleotide and amino-acid characteristics, it may be that amino acids, peptides, and finally proteins initially were cofactors for ribozymes.
One of the challenges in studying abiogenesis is that the system of reproduction and metabolism utilized by all extant life involves three distinct types of interdependent macromolecules (DNA, RNA, and proteins), none of which can function and reproduce without the others, the classic chicken-and-egg paradox. This suggests that life could not have arisen in its current form, which has led researchers to hypothesize mechanisms whereby the current system might have arisen from a simpler precursor system. American molecular biologist Alexander Rich was the first to posit a coherent hypothesis on the origin of nucleotides as precursors of life. In an article he contributed to a volume issued in honor of Nobel-laureate physiologist Albert Szent-Györgyi, he explained that the primitive Earth's environment could have produced RNA molecules (polynucleotide monomers) that eventually acquired enzymatic and self-replicating functions.
Other mentions of RNA as a primordial molecule can be found in papers by Francis Crick and Leslie Orgel, as well as in Carl Woese's 1967 book The Genetic Code. Hans Kuhn in 1972 laid out a possible process by which the modern genetic system might have arisen from a nucleotide-based precursor, and this led Harold White in 1976 to observe that many of the cofactors essential for enzymatic function are either nucleotides or could have been derived from nucleotides. He proposed a scenario whereby the critical electrochemistry of enzymatic reactions would have necessitated retention of the specific nucleotide moieties of the original RNA-based enzymes carrying out the reactions, while the remaining structural elements of the enzymes were gradually replaced by protein, until all that remained of the original RNAs were these nucleotide cofactors, "fossils of nucleic acid enzymes".
The properties of RNA make the idea of the RNA world hypothesis conceptually plausible, though its general acceptance as an explanation for the origin of life requires further evidence. RNA is known to form efficient catalysts, and its similarity to DNA makes clear its ability to store information. Opinions differ, however, as to whether RNA constituted the first autonomous self-replicating system or was a derivative of a still-earlier system. One version of the hypothesis is that a different type of nucleic acid, termed pre-RNA, was the first one to emerge as a self-reproducing molecule, to be replaced by RNA only later. On the other hand, the discovery in 2009 that activated pyrimidine ribonucleotides can be synthesized under plausible prebiotic conditions suggests that it is premature to dismiss the RNA-first scenarios. Suggestions for 'simple' pre-RNA nucleic acids have included peptide nucleic acid (PNA), threose nucleic acid (TNA) or glycol nucleic acid (GNA). Despite their structural simplicity and possession of properties comparable with RNA, the chemically plausible generation of "simpler" nucleic acids under prebiotic conditions has yet to be demonstrated.
In the 1980s, RNA structures capable of self-processing were discovered, with the RNA moiety of ribonuclease P acting as its catalytic subunit. These catalytic RNAs – referred to as RNA enzymes, or ribozymes – are found in today's DNA-based life and could be examples of living fossils. Ribozymes play vital roles, such as that of the ribosome. The large subunit of the ribosome includes an rRNA responsible for the peptide bond-forming peptidyl transferase activity of protein synthesis. Many other ribozyme activities exist; for example, the hammerhead ribozyme performs self-cleavage and an RNA polymerase ribozyme can synthesize a short RNA strand from a primed RNA template.
RNA world
The RNA world is a hypothetical stage in the evolutionary history of life on Earth in which self-replicating RNA molecules proliferated before the evolution of DNA and proteins. The term also refers to the hypothesis that posits the existence of this stage. Alexander Rich first proposed the concept of the RNA world in 1962, and Walter Gilbert coined the term in 1986.
Among the characteristics of RNA that suggest its original prominence are that:
Although alternative chemical paths to life have been proposed, and RNA-based life may not have been the first life to exist, the RNA world hypothesis seems to be the most favored abiogenesis paradigm. However, even proponents agree that there is still not conclusive evidence to completely falsify other paradigms and hypotheses. Regardless of its plausibility in a prebiotic scenario, the RNA world can serve as a model system for studying the origin of life.
If the RNA world existed, it was probably followed by an age characterized by the evolution of ribonucleoproteins (RNP world), which in turn ushered in the era of DNA and longer proteins. DNA has greater stability and durability than RNA, which may explain why it became the predominant information storage molecule. Protein enzymes may have replaced RNA-based ribozymes as biocatalysts because the greater abundance and diversity of the monomers of which they are built makes them more versatile. As some cofactors contain both nucleotide and amino-acid characteristics, it may be that amino acids, peptides, and finally proteins initially were cofactors for ribozymes.
One of the challenges in studying abiogenesis is that the system of reproduction and metabolism utilized by all extant life involves three distinct types of interdependent macromolecules (DNA, RNA, and proteins), none of which can function and reproduce without the others, the classic chicken-and-egg paradox. This suggests that life could not have arisen in its current form, which has led researchers to hypothesize mechanisms whereby the current system might have arisen from a simpler precursor system. American molecular biologist Alexander Rich was the first to posit a coherent hypothesis on the origin of nucleotides as precursors of life. In an article he contributed to a volume issued in honor of Nobel-laureate physiologist Albert Szent-Györgyi, he explained that the primitive Earth's environment could have produced RNA molecules (polynucleotide monomers) that eventually acquired enzymatic and self-replicating functions.
Other mentions of RNA as a primordial molecule can be found in papers by Francis Crick and Leslie Orgel, as well as in Carl Woese's 1967 book The Genetic Code. Hans Kuhn in 1972 laid out a possible process by which the modern genetic system might have arisen from a nucleotide-based precursor, and this led Harold White in 1976 to observe that many of the cofactors essential for enzymatic function are either nucleotides or could have been derived from nucleotides. He proposed a scenario whereby the critical electrochemistry of enzymatic reactions would have necessitated retention of the specific nucleotide moieties of the original RNA-based enzymes carrying out the reactions, while the remaining structural elements of the enzymes were gradually replaced by protein, until all that remained of the original RNAs were these nucleotide cofactors, "fossils of nucleic acid enzymes".
The properties of RNA make the idea of the RNA world hypothesis conceptually plausible, though its general acceptance as an explanation for the origin of life requires further evidence. RNA is known to form efficient catalysts, and its similarity to DNA makes clear its ability to store information. Opinions differ, however, as to whether RNA constituted the first autonomous self-replicating system or was a derivative of a still-earlier system. One version of the hypothesis is that a different type of nucleic acid, termed pre-RNA, was the first one to emerge as a self-reproducing molecule, to be replaced by RNA only later. On the other hand, the discovery in 2009 that activated pyrimidine ribonucleotides can be synthesized under plausible prebiotic conditions suggests that it is premature to dismiss the RNA-first scenarios. Suggestions for 'simple' pre-RNA nucleic acids have included peptide nucleic acid (PNA), threose nucleic acid (TNA) or glycol nucleic acid (GNA). Despite their structural simplicity and possession of properties comparable with RNA, the chemically plausible generation of "simpler" nucleic acids under prebiotic conditions has yet to be demonstrated.
In the 1980s, RNA structures capable of self-processing were discovered, with the RNA moiety of ribonuclease P acting as its catalytic subunit. These catalytic RNAs – referred to as RNA enzymes, or ribozymes – are found in today's DNA-based life and could be examples of living fossils. Ribozymes play vital roles, such as that of the ribosome. The large subunit of the ribosome includes an rRNA responsible for the peptide bond-forming peptidyl transferase activity of protein synthesis. Many other ribozyme activities exist; for example, the hammerhead ribozyme performs self-cleavage and an RNA polymerase ribozyme can synthesize a short RNA strand from a primed RNA template.
Recent media
Recent media