
Community hub
Capitulum of the humerus
View on WikipediaThis article includes a list of general references, but it lacks sufficient corresponding inline citations. (December 2013) |
| Capitulum of the humerus | |
|---|---|
 Left humerus seen from front (capitulum visible at bottom right) | |
 Left humerus seen from front (part of the appendicular skeleton) | |
| Details | |
| Identifiers | |
| Latin | capitulum humeri |
| TA98 | A02.4.04.022 |
| TA2 | 1202 |
| FMA | 23373 |
| Anatomical terms of bone | |
In human anatomy of the arm, the capitulum of the humerus is a smooth, rounded eminence on the lateral portion of the distal articular surface of the humerus. It articulates with the cup-shaped depression on the head of the radius, and is limited to the front and lower part of the bone.
In non-human tetrapods, the name capitellum is generally used, with "capitulum" limited to the anteroventral articular facet of the rib (in archosauromorphs).
Lepidosauromorpha
[edit]Lepidosaurs show a distinct capitellum and trochlea on the centre of the ventral (anterior in upright taxa) surface of the humerus at the distal end.
Archosauromorpha
[edit]In non-avian archosaurs, including crocodiles, the capitellum and the trochlea are no longer bordered by distinct etc.- and entepicondyles respectively, and the distal humerus consists two gently expanded condyles, one lateral and one medial, separated by a shallow groove and a supinator process. Romer (1976) homologizes the capitellum in Archosauromorphs with the groove separating the medial and lateral condyles.
In birds, where forelimb anatomy has an adaptation for flight, its functional if not[1] ontogenetic equivalent is the dorsal condyle of the humerus.
Additional images
[edit]-
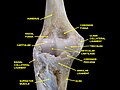 Elbow joint. Deep dissection. Anterior view.
Elbow joint. Deep dissection. Anterior view. -
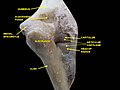 Elbow joint. Deep dissection. Posterior view.
Elbow joint. Deep dissection. Posterior view. -
 Elbow joint. Deep dissection. Posterior view.
Elbow joint. Deep dissection. Posterior view.
.jpg)


References
[edit]- ^ "Evolution of Birds". The George Washington University.
![]() This article incorporates text in the public domain from page 212 of the 20th edition of Gray's Anatomy (1918)
This article incorporates text in the public domain from page 212 of the 20th edition of Gray's Anatomy (1918)
- Romer, A.S. 1976 Osteology of the reptiles. University of Chicago Press, Chicago.
External links
[edit]- Anatomy figure: 07:02-05 at Human Anatomy Online, SUNY Downstate Medical Center
- BiowebUW, cached at archive.org