
Community hub
Recent from talks
Contribute something to knowledge base
Content stats: 0 posts, 0 articles, 1 media, 0 notes
Members stats: 0 subscribers, 0 contributors, 0 moderators, 0 supporters
Subscribers
Supporters
Contributors
Moderators
Hub AI
Cartilage AI simulator
(@Cartilage_simulator)
Hub AI
Cartilage AI simulator
(@Cartilage_simulator)
Cartilage
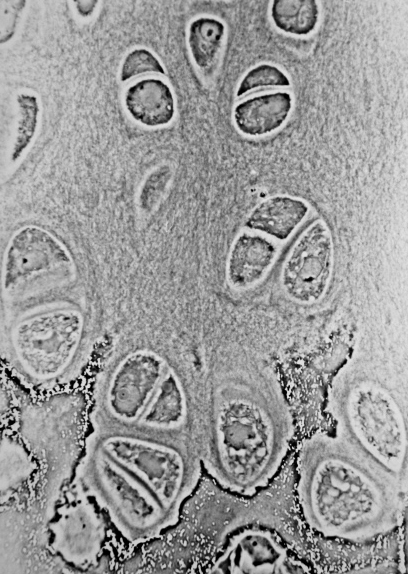
Cartilage is a resilient and smooth type of connective tissue. Semi-transparent and non-porous, it is usually covered by a tough and fibrous membrane called perichondrium. In tetrapods, it covers and protects the ends of long bones at the joints as articular cartilage, and is a structural component of many body parts including the rib cage, the neck and the bronchial tubes, and the intervertebral discs. In other taxa, such as chondrichthyans and cyclostomes, it constitutes a much greater proportion of the skeleton. It is not as hard and rigid as bone, but it is much stiffer and much less flexible than muscle or tendon. The matrix of cartilage is made up of glycosaminoglycans, proteoglycans, collagen fibers and, sometimes, elastin. It usually grows quicker than bone.
Because of its rigidity, cartilage often serves the purpose of holding tubes open in the body. Examples include the rings of the trachea, such as the cricoid cartilage and carina.
Cartilage is composed of specialized cells called chondrocytes that produce a large amount of collagenous extracellular matrix, abundant ground substance that is rich in proteoglycan and elastin fibers. Cartilage is classified into three types — elastic cartilage, hyaline cartilage, and fibrocartilage — which differ in their relative amounts of collagen and proteoglycan.
As cartilage does not contain blood vessels or nerves, it is insensitive. However, some fibrocartilage such as the meniscus of the knee has partial blood supply. Nutrition is supplied to the chondrocytes by diffusion. The compression of the articular cartilage or flexion of the elastic cartilage generates fluid flow, which assists the diffusion of nutrients to the chondrocytes. Compared to other connective tissues, cartilage has a very slow turnover of its extracellular matrix and is documented to repair at only a very slow rate relative to other tissues.
In embryogenesis, the skeletal system is derived from the mesoderm germ layer. Chondrification (also known as chondrogenesis) is the process by which cartilage is formed from condensed mesenchyme tissue, which differentiates into chondroblasts and begins secreting the molecules (aggrecan and collagen type II) that form the extracellular matrix. In all vertebrates, cartilage is the main skeletal tissue in early ontogenetic stages; in osteichthyans, many cartilaginous elements subsequently ossify through endochondral and perichondral ossification.
Following the initial chondrification that occurs during embryogenesis, cartilage growth consists mostly of the maturing of immature cartilage to a more mature state. The division of cells within cartilage occurs very slowly, and thus growth in cartilage is usually not based on an increase in size or mass of the cartilage itself. It has been identified that non-coding RNAs (e.g. miRNAs and long non-coding RNAs) as the most important epigenetic modulators can affect the chondrogenesis. This also justifies the non-coding RNAs' contribution in various cartilage-dependent pathological conditions such as arthritis, and so on.
The articular cartilage function is dependent on the molecular composition of the extracellular matrix (ECM). The ECM consists mainly of proteoglycan and collagens. The main proteoglycan in cartilage is aggrecan, which, as its name suggests, forms large aggregates with hyaluronan and with itself. These aggregates are negatively charged and hold water in the tissue. The collagen, mostly collagen type II, constrains the proteoglycans. The ECM responds to tensile and compressive forces that are experienced by the cartilage. Cartilage growth thus refers to the matrix deposition, but can also refer to both the growth and remodeling of the extracellular matrix. Due to the great stress on the patellofemoral joint during resisted knee extension, the articular cartilage of the patella is among the thickest in the human body. The ECM of articular cartilage is classified into three regions: the pericellular matrix, the territorial matrix, and the interterritorial matrix.
The mechanical properties of articular cartilage in load-bearing joints such as the knee and hip have been studied extensively at macro, micro, and nano-scales. These mechanical properties include the response of cartilage in frictional, compressive, shear and tensile loading. Cartilage is resilient and displays viscoelastic properties.
Cartilage
Cartilage is a resilient and smooth type of connective tissue. Semi-transparent and non-porous, it is usually covered by a tough and fibrous membrane called perichondrium. In tetrapods, it covers and protects the ends of long bones at the joints as articular cartilage, and is a structural component of many body parts including the rib cage, the neck and the bronchial tubes, and the intervertebral discs. In other taxa, such as chondrichthyans and cyclostomes, it constitutes a much greater proportion of the skeleton. It is not as hard and rigid as bone, but it is much stiffer and much less flexible than muscle or tendon. The matrix of cartilage is made up of glycosaminoglycans, proteoglycans, collagen fibers and, sometimes, elastin. It usually grows quicker than bone.
Because of its rigidity, cartilage often serves the purpose of holding tubes open in the body. Examples include the rings of the trachea, such as the cricoid cartilage and carina.
Cartilage is composed of specialized cells called chondrocytes that produce a large amount of collagenous extracellular matrix, abundant ground substance that is rich in proteoglycan and elastin fibers. Cartilage is classified into three types — elastic cartilage, hyaline cartilage, and fibrocartilage — which differ in their relative amounts of collagen and proteoglycan.
As cartilage does not contain blood vessels or nerves, it is insensitive. However, some fibrocartilage such as the meniscus of the knee has partial blood supply. Nutrition is supplied to the chondrocytes by diffusion. The compression of the articular cartilage or flexion of the elastic cartilage generates fluid flow, which assists the diffusion of nutrients to the chondrocytes. Compared to other connective tissues, cartilage has a very slow turnover of its extracellular matrix and is documented to repair at only a very slow rate relative to other tissues.
In embryogenesis, the skeletal system is derived from the mesoderm germ layer. Chondrification (also known as chondrogenesis) is the process by which cartilage is formed from condensed mesenchyme tissue, which differentiates into chondroblasts and begins secreting the molecules (aggrecan and collagen type II) that form the extracellular matrix. In all vertebrates, cartilage is the main skeletal tissue in early ontogenetic stages; in osteichthyans, many cartilaginous elements subsequently ossify through endochondral and perichondral ossification.
Following the initial chondrification that occurs during embryogenesis, cartilage growth consists mostly of the maturing of immature cartilage to a more mature state. The division of cells within cartilage occurs very slowly, and thus growth in cartilage is usually not based on an increase in size or mass of the cartilage itself. It has been identified that non-coding RNAs (e.g. miRNAs and long non-coding RNAs) as the most important epigenetic modulators can affect the chondrogenesis. This also justifies the non-coding RNAs' contribution in various cartilage-dependent pathological conditions such as arthritis, and so on.
The articular cartilage function is dependent on the molecular composition of the extracellular matrix (ECM). The ECM consists mainly of proteoglycan and collagens. The main proteoglycan in cartilage is aggrecan, which, as its name suggests, forms large aggregates with hyaluronan and with itself. These aggregates are negatively charged and hold water in the tissue. The collagen, mostly collagen type II, constrains the proteoglycans. The ECM responds to tensile and compressive forces that are experienced by the cartilage. Cartilage growth thus refers to the matrix deposition, but can also refer to both the growth and remodeling of the extracellular matrix. Due to the great stress on the patellofemoral joint during resisted knee extension, the articular cartilage of the patella is among the thickest in the human body. The ECM of articular cartilage is classified into three regions: the pericellular matrix, the territorial matrix, and the interterritorial matrix.
The mechanical properties of articular cartilage in load-bearing joints such as the knee and hip have been studied extensively at macro, micro, and nano-scales. These mechanical properties include the response of cartilage in frictional, compressive, shear and tensile loading. Cartilage is resilient and displays viscoelastic properties.
Recent media
Recent media