Recent from talks
Motor protein
Knowledge base stats:
Talk channels stats:
Members stats:
Motor protein
Motor proteins are a class of molecular motors that can move along the cytoskeleton of cells. They do this by converting chemical energy into mechanical work by the hydrolysis of ATP.
Motor proteins are the driving force behind most active transport of proteins and vesicles in the cytoplasm. Kinesins and cytoplasmic dyneins play essential roles in intracellular transport such as axonal transport and in the formation of the spindle apparatus and the separation of the chromosomes during mitosis and meiosis. Axonemal dynein, found in cilia and flagella, is crucial to cell motility in spermatozoa, and fluid transport in trachea.[citation needed] The muscle protein myosin "motors" the contraction of muscle fibers in animals.
The importance of motor proteins in cells becomes evident when they fail to fulfill their function. For example, kinesin deficiencies have been identified as the cause for Charcot-Marie-Tooth disease and some kidney diseases. Dynein deficiencies can lead to chronic infections of the respiratory tract as cilia fail to function without dynein. Numerous myosin deficiencies are related to disease states and genetic syndromes. Because myosin II is essential for muscle contraction, defects in muscular myosin predictably cause myopathies. Myosin is necessary in the process of hearing because of its role in the growth of stereocilia so defects in myosin protein structure can lead to Usher syndrome and non-syndromic deafness.
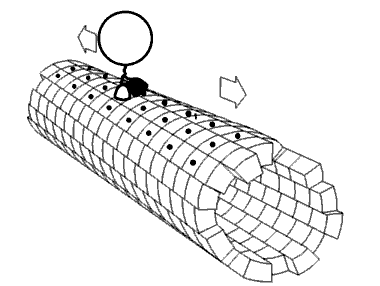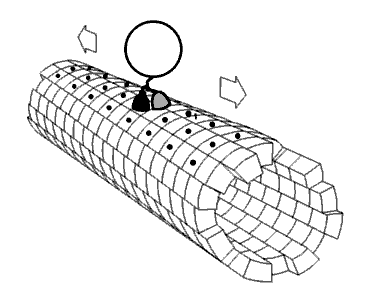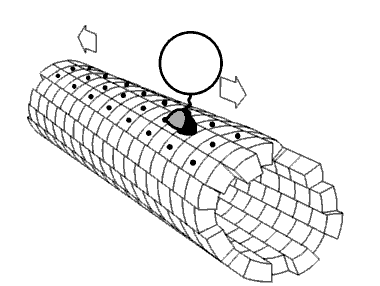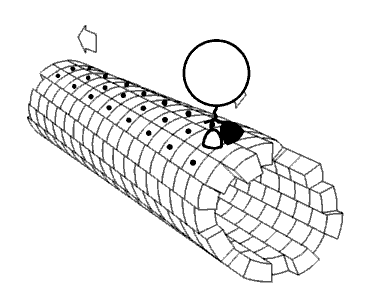
Motor proteins utilizing the cytoskeleton for movement fall into two categories based on their substrate: microfilaments or microtubules. Actin-based motor proteins (myosin) move along microfilaments through interaction with actin, and microtubule motors (dynein and kinesin) move along microtubules through interaction with tubulin.[citation needed]
There are two basic types of microtubule motors: plus-end motors and minus-end motors, depending on the direction in which they "walk" along the microtubule cables within the cell.
Myosins are a superfamily of actin motor proteins that convert chemical energy in the form of ATP to mechanical energy, thus generating force and movement. The first identified myosin, myosin II, is responsible for generating muscle contraction. Myosin II is an elongated protein that is formed from two heavy chains with motor heads and two light chains. Each myosin head contains actin and ATP binding site. The myosin heads bind and hydrolyze ATP, which provides the energy to walk toward the plus end of an actin filament. Myosin II are also vital in the process of cell division. For example, non-muscle myosin II bipolar thick filaments provide the force of contraction needed to divide the cell into two daughter cells during cytokinesis. In addition to myosin II, many other myosin types are responsible for variety of movement of non-muscle cells. For example, myosin is involved in intracellular organization and the protrusion of actin-rich structures at the cell surface. Myosin V is involved in vesicle and organelle transport. Myosin XI is involved in cytoplasmic streaming, wherein movement along microfilament networks in the cell allows organelles and cytoplasm to stream in a particular direction. Eighteen different classes of myosins are known.
Genomic representation of myosin motors:
Kinesins are a superfamily of related motor proteins that use a microtubule track in anterograde movement. They are vital to spindle formation in mitotic and meiotic chromosome separation during cell division and are also responsible for shuttling mitochondria, Golgi bodies, and vesicles within eukaryotic cells. Kinesins have two heavy chains and two light chains per active motor. The two globular head motor domains in heavy chains can convert the chemical energy of ATP hydrolysis into mechanical work to move along microtubules. The direction in which cargo is transported can be towards the plus-end or the minus-end, depending on the type of kinesin. In general, kinesins with N-terminal motor domains move their cargo towards the plus ends of microtubules located at the cell periphery, while kinesins with C-terminal motor domains move cargo towards the minus ends of microtubules located at the nucleus. Fourteen distinct kinesin families are known, with some additional kinesin-like proteins that cannot be classified into these families.
Hub AI
Motor protein AI simulator
(@Motor protein_simulator)
Motor protein
Motor proteins are a class of molecular motors that can move along the cytoskeleton of cells. They do this by converting chemical energy into mechanical work by the hydrolysis of ATP.
Motor proteins are the driving force behind most active transport of proteins and vesicles in the cytoplasm. Kinesins and cytoplasmic dyneins play essential roles in intracellular transport such as axonal transport and in the formation of the spindle apparatus and the separation of the chromosomes during mitosis and meiosis. Axonemal dynein, found in cilia and flagella, is crucial to cell motility in spermatozoa, and fluid transport in trachea.[citation needed] The muscle protein myosin "motors" the contraction of muscle fibers in animals.
The importance of motor proteins in cells becomes evident when they fail to fulfill their function. For example, kinesin deficiencies have been identified as the cause for Charcot-Marie-Tooth disease and some kidney diseases. Dynein deficiencies can lead to chronic infections of the respiratory tract as cilia fail to function without dynein. Numerous myosin deficiencies are related to disease states and genetic syndromes. Because myosin II is essential for muscle contraction, defects in muscular myosin predictably cause myopathies. Myosin is necessary in the process of hearing because of its role in the growth of stereocilia so defects in myosin protein structure can lead to Usher syndrome and non-syndromic deafness.
Motor proteins utilizing the cytoskeleton for movement fall into two categories based on their substrate: microfilaments or microtubules. Actin-based motor proteins (myosin) move along microfilaments through interaction with actin, and microtubule motors (dynein and kinesin) move along microtubules through interaction with tubulin.[citation needed]
There are two basic types of microtubule motors: plus-end motors and minus-end motors, depending on the direction in which they "walk" along the microtubule cables within the cell.
Myosins are a superfamily of actin motor proteins that convert chemical energy in the form of ATP to mechanical energy, thus generating force and movement. The first identified myosin, myosin II, is responsible for generating muscle contraction. Myosin II is an elongated protein that is formed from two heavy chains with motor heads and two light chains. Each myosin head contains actin and ATP binding site. The myosin heads bind and hydrolyze ATP, which provides the energy to walk toward the plus end of an actin filament. Myosin II are also vital in the process of cell division. For example, non-muscle myosin II bipolar thick filaments provide the force of contraction needed to divide the cell into two daughter cells during cytokinesis. In addition to myosin II, many other myosin types are responsible for variety of movement of non-muscle cells. For example, myosin is involved in intracellular organization and the protrusion of actin-rich structures at the cell surface. Myosin V is involved in vesicle and organelle transport. Myosin XI is involved in cytoplasmic streaming, wherein movement along microfilament networks in the cell allows organelles and cytoplasm to stream in a particular direction. Eighteen different classes of myosins are known.
Genomic representation of myosin motors:
Kinesins are a superfamily of related motor proteins that use a microtubule track in anterograde movement. They are vital to spindle formation in mitotic and meiotic chromosome separation during cell division and are also responsible for shuttling mitochondria, Golgi bodies, and vesicles within eukaryotic cells. Kinesins have two heavy chains and two light chains per active motor. The two globular head motor domains in heavy chains can convert the chemical energy of ATP hydrolysis into mechanical work to move along microtubules. The direction in which cargo is transported can be towards the plus-end or the minus-end, depending on the type of kinesin. In general, kinesins with N-terminal motor domains move their cargo towards the plus ends of microtubules located at the cell periphery, while kinesins with C-terminal motor domains move cargo towards the minus ends of microtubules located at the nucleus. Fourteen distinct kinesin families are known, with some additional kinesin-like proteins that cannot be classified into these families.
Recent media