Recent from talks
Asiavorator
Knowledge base stats:
Talk channels stats:
Members stats:
Asiavorator
Asiavorator (meaning "Asian devourer") is an extinct genus of civet-like carnivoran belonging in the family Stenoplesictidae. It was endemic to Asia and lived during the Eocene and Oligocene epochs.
The teeth of Asiavorator suggest that it was omnivorous or more precisely, ranged from hypercarnivorous to mesocarnivorous.
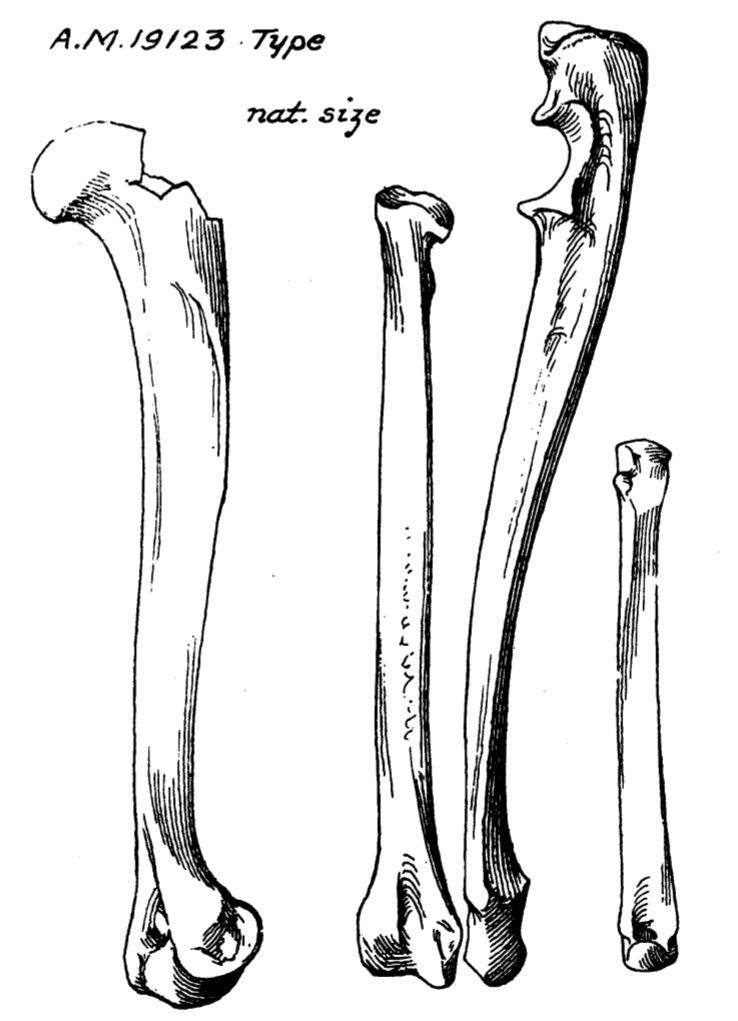
The first remains of Asiavorator to be found were collected in the 1922 field season of the Central Asiatic Expeditions near the Loh campsite in Övörkhangai Province, Mongolia. This locality is part of the Hsanda Gol Formation. The specimens, designated AMNH 19123, included limb bones and lower teeth. Matthew and Granger (1924) described AMNH 19123 as the type specimen of a new carnivoran species they named Palaeoprionodon gracilis.
The genus Asiavorator was erected by Spassov and Lange-Badré in 1995 as a monotypic genus for their new species A. altidens, with the type specimen of A. altidens being a mandible (FM 487-95) from the Hsanda Gol Formation. Dashzeveg (1996) described a new species of stenoplesictid, Stenoplesictis simplex, based on a mandible (PSS 27-25) from the Ergilin Dzo Formation of Mongolia. In 1998, Hunt reassigned S. simplex to the genus Shandgolictis, renaming it Shandgolictis simplex and assigning it to Aeluroidea.
Later authors found that Asiavorator altidens and Palaeoprionodon gracilis were synonymous and represent a distinct genus, thus the two were synonymized as Asiavorator gracilis, retaining the specific name of the latter and the generic name of the former. A re-examination by Egi et al. (2016) found that the tooth measurements of PSS 27-25 are not notably different from those of AMNH 19123, thus concluding that Stenoplesictis simplex and Shandgolictis simplex are junior synonyms of Asiavorator gracilis. Currently, A. gracilis is the only accepted species in the genus.
Using the carnivoran regression on the specimen PSS 21-25, Asiavorator has been estimated to have a body mass of 3.6 to 5.6 kg. This is larger than Alagtsavbaatar, a feliform known to have been sympatric with Asiavorator, whose body mass has been estimated at 2.6 to 3.6 kg.
Like many other carnivorous mammals, Asiavorator has long and sharp-pointed canine teeth, presumably used in killing prey. The upper and lower canines were approximately equal in length. The carnassial resembles that of a cat, being compressed and possessing a vestigial heel and reduced metaconid. The first upper molar is very elongated, measuring 10 mm long and 4.5 mm wide in the specimen PSS 27-25. An obtuse angle is formed by the shearing edges of the protoconid and paraconid, while the well-developed metaconid is placed against the internal posterior side of the protoconid. The base of the crown has a cingulum on the external side. The second molar is bunodont, and possesses two roots and a flattened trigonid of three low cusps and a trenchant heel. The fourth premolar is large and compressed, similar to the condition seen in domestic cats.
Asiavorator had a well-developed masseteric fossa and a thin mandible. The mandibular corpus has a prominent lower edge below the molars. Below the first molar, the mandibular ramus of the specimen PSS 27-25 measures 13.4 mm in height and 5.6 mm in width.
Hub AI
Asiavorator AI simulator
(@Asiavorator_simulator)
Asiavorator
Asiavorator (meaning "Asian devourer") is an extinct genus of civet-like carnivoran belonging in the family Stenoplesictidae. It was endemic to Asia and lived during the Eocene and Oligocene epochs.
The teeth of Asiavorator suggest that it was omnivorous or more precisely, ranged from hypercarnivorous to mesocarnivorous.
The first remains of Asiavorator to be found were collected in the 1922 field season of the Central Asiatic Expeditions near the Loh campsite in Övörkhangai Province, Mongolia. This locality is part of the Hsanda Gol Formation. The specimens, designated AMNH 19123, included limb bones and lower teeth. Matthew and Granger (1924) described AMNH 19123 as the type specimen of a new carnivoran species they named Palaeoprionodon gracilis.
The genus Asiavorator was erected by Spassov and Lange-Badré in 1995 as a monotypic genus for their new species A. altidens, with the type specimen of A. altidens being a mandible (FM 487-95) from the Hsanda Gol Formation. Dashzeveg (1996) described a new species of stenoplesictid, Stenoplesictis simplex, based on a mandible (PSS 27-25) from the Ergilin Dzo Formation of Mongolia. In 1998, Hunt reassigned S. simplex to the genus Shandgolictis, renaming it Shandgolictis simplex and assigning it to Aeluroidea.
Later authors found that Asiavorator altidens and Palaeoprionodon gracilis were synonymous and represent a distinct genus, thus the two were synonymized as Asiavorator gracilis, retaining the specific name of the latter and the generic name of the former. A re-examination by Egi et al. (2016) found that the tooth measurements of PSS 27-25 are not notably different from those of AMNH 19123, thus concluding that Stenoplesictis simplex and Shandgolictis simplex are junior synonyms of Asiavorator gracilis. Currently, A. gracilis is the only accepted species in the genus.
Using the carnivoran regression on the specimen PSS 21-25, Asiavorator has been estimated to have a body mass of 3.6 to 5.6 kg. This is larger than Alagtsavbaatar, a feliform known to have been sympatric with Asiavorator, whose body mass has been estimated at 2.6 to 3.6 kg.
Like many other carnivorous mammals, Asiavorator has long and sharp-pointed canine teeth, presumably used in killing prey. The upper and lower canines were approximately equal in length. The carnassial resembles that of a cat, being compressed and possessing a vestigial heel and reduced metaconid. The first upper molar is very elongated, measuring 10 mm long and 4.5 mm wide in the specimen PSS 27-25. An obtuse angle is formed by the shearing edges of the protoconid and paraconid, while the well-developed metaconid is placed against the internal posterior side of the protoconid. The base of the crown has a cingulum on the external side. The second molar is bunodont, and possesses two roots and a flattened trigonid of three low cusps and a trenchant heel. The fourth premolar is large and compressed, similar to the condition seen in domestic cats.
Asiavorator had a well-developed masseteric fossa and a thin mandible. The mandibular corpus has a prominent lower edge below the molars. Below the first molar, the mandibular ramus of the specimen PSS 27-25 measures 13.4 mm in height and 5.6 mm in width.
Recent media