Community hub
Factor XI
View on Wikipedia
| F11 | |||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 | |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| Identifiers | |||||||||||||||||||||||||||||||||||||||||||||||||||
| Aliases | F11, FXI, coagulation factor XI, PTA, factor XI | ||||||||||||||||||||||||||||||||||||||||||||||||||
| External IDs | OMIM: 264900; MGI: 99481; HomoloGene: 86654; GeneCards: F11; OMA:F11 - orthologs | ||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| Wikidata | |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||





Factor XI, or plasma thromboplastin antecedent, is the zymogen form of factor XIa, one of the enzymes involved in coagulation. Like many other coagulation factors, it is a serine protease. In humans, factor XI is encoded by F11 gene.[5][6][7][8]
Function
[edit]Factor XI (FXI) is produced by the liver and circulates as a homo-dimer in its inactive form.[9] The plasma half-life of FXI is approximately 52 hours. The zymogen factor is activated into factor XIa by factor XIIa (FXIIa), thrombin, and FXIa itself; due to its activation by FXIIa, FXI is a member of the "contact pathway" (which includes HMWK, prekallikrein, factor XII, factor XI, and factor IX).[10]
Factor XIa activates factor IX by selectively cleaving arg-ala and arg-val peptide bonds. Factor IXa, in turn, forms a complex with Factor VIIIa (FIXa-FVIIIa) and activates factor X.
Physiological inhibitors of factor XIa include protein Z-dependent protease inhibitor (ZPI, a member of the serine protease inhibitor/serpin class of proteins), which is independent of protein Z (its action on factor X, however, is protein Z-dependent, hence its name).
Structure
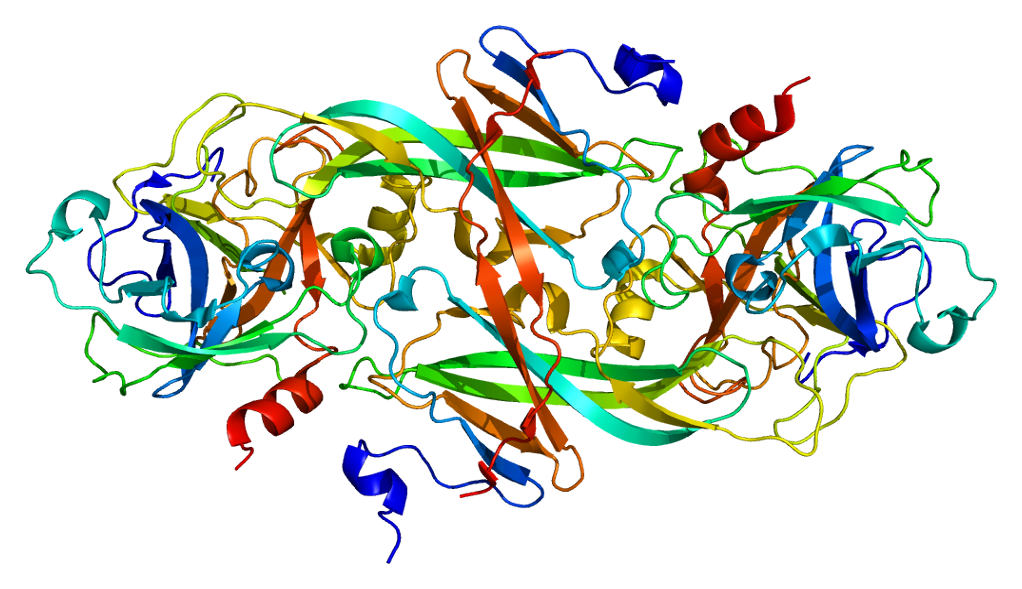
[edit]Although synthesized as a single polypeptide chain, FXI circulates as a homodimer. Every chain has a relative molecular mass of approximately 80000. Typical plasma concentrations of FXI are 5 μg/mL, corresponding to a plasma concentration (of FXI dimers) of approximately 30 nM. The FXI gene is 23kb in length, has 15 exons, and is found on chromosome 4q32-35.[6][7]
Factor XI consists of four apple domains, that create a disk-like platform around the base of a fifth, catalytic serine protease domain. One contains a binding site for thrombin, another for high molecular weight kininogen, a third one for factor IX, heparin and glycoprotein Ib and the fourth is implicated in forming the factor XI homodimer, including a cysteine residue that creates a disulfide bond.
In the homodimer, the apple domains create two disk-like platforms connected together at an angle, with the catalytic domains sticking out at each side of the dimer.
Activation by thrombin or factor XIIa is achieved by cleavage of Arg369-Ile370 peptide bonds on both subunits of the dimer. This results in a partial detachment of the catalytic domain from the disk-like apple domains, still linked to the fourth domain with a disulfide bond, but now farther from the third domain. This is thought that this exposes the factor IX binding site of the third apple domain, allowing factor XI's protease activity on it. [11]
Role in disease
[edit]Deficiency of factor XI causes the rare hemophilia C; this mainly occurs in Ashkenazi Jews and is believed to affect approximately 8% of that population. Less commonly, hemophilia C can be found in Jews of Iraqi ancestry and in Israeli Arabs. The condition has been described in other populations at around 1% of cases. There is little spontaneous bleeding, but surgical procedures may cause excessive blood loss, and prophylaxis is required.[12]
Low levels of factor XI also occur in many other disease states, including Noonan syndrome.
High levels of factor XI have been implicated in thrombosis, although it is uncertain what determines these levels and how serious the procoagulant state is.
Inhibition
[edit]Pharmacological inhibitors of factor XI that are under clinical development but not yet approved for treatment as of May 2022[update] include the oral factor XIa inhibitors Asundexian (BAY 2433334)[13] and Milvexian[14] as well as the monoclonal anti-factor XI antibody abelacimab (MAA868). The idea behind producing such an inhibitor is that XI is mostly involved in intrinsic/contact activation pathway,[15] which plays a bigger role in thrombosis as opposed to hemostasis, so targeting it may reduce clotting risks without a corresponding increase in bleeding.[16] An abelacimab trial appears to have indeed produced this result.[17]
See also
[edit]- Contact activation pathway (also known as the intrinsic pathway)
- Tissue factor pathway (also known as the extrinsic pathway)
References
[edit]- ^ a b c GRCh38: Ensembl release 89: ENSG00000088926 – Ensembl, May 2017
- ^ a b c GRCm38: Ensembl release 89: ENSMUSG00000031645 – Ensembl, May 2017
- ^ "Human PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- ^ "Mouse PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- ^ Fujikawa K, Chung DW, Hendrickson LE, Davie EW (May 1986). "Amino acid sequence of human factor XI, a blood coagulation factor with four tandem repeats that are highly homologous with plasma prekallikrein". Biochemistry. 25 (9): 2417–2424. doi:10.1021/bi00357a018. PMID 3636155.
- ^ a b Asakai R, Davie EW, Chung DW (Nov 1987). "Organization of the gene for human factor XI". Biochemistry. 26 (23): 7221–7228. doi:10.1021/bi00397a004. PMID 2827746.
- ^ a b Kato A, Asakai R, Davie EW, Aoki N (1989). "Factor XI gene (F11) is located on the distal end of the long arm of human chromosome 4". Cytogenetics and Cell Genetics. 52 (1–2): 77–78. doi:10.1159/000132844. PMID 2612218.
- ^ Buetow KH, Shiang R, Yang P, Nakamura Y, Lathrop GM, White R, et al. (May 1991). "A detailed multipoint map of human chromosome 4 provides evidence for linkage heterogeneity and position-specific recombination rates". American Journal of Human Genetics. 48 (5): 911–925. PMC 1683054. PMID 1673289.
- ^ Wu W, Sinha D, Shikov S, Yip CK, Walz T, Billings PC, et al. (Jul 2008). "Factor XI homodimer structure is essential for normal proteolytic activation by factor XIIa, thrombin, and factor XIa". The Journal of Biological Chemistry. 283 (27): 18655–18664. doi:10.1074/jbc.M802275200. PMC 2441546. PMID 18441012.
- ^ Walsh PN (Jul 2001). "Roles of platelets and factor XI in the initiation of blood coagulation by thrombin". Thrombosis and Haemostasis. 86 (1): 75–82. doi:10.1055/s-0037-1616203. PMID 11487044. S2CID 32572142. Archived from the original on 2016-04-16. Retrieved 2009-01-07.
- ^ Emsley J, McEwan PA, Gailani D (Apr 2010). "Structure and function of factor XI". Blood. 115 (13): 2569–2577. doi:10.1182/blood-2009-09-199182. PMC 4828079. PMID 20110423.
- ^ Bolton-Maggs PH (Jun 1996). "Factor XI deficiency". Bailliere's Clinical Haematology. 9 (2): 355–368. doi:10.1016/S0950-3536(96)80068-0. PMID 8800510.
- ^ Clinical trial number NCT04304508 for "Study to Gather Information About Proper Dosing and Safety of the Oral FXIa Inhibitor BAY 2433334 in Patients Following a Recent Non Cardioembolic Ischemic Stroke Which Occurs When a Blood Clot Has Formed Somewhere in the Human Body (But Not in the Heart) Travelled to the Brain. (PACIFIC-STROKE)" at ClinicalTrials.gov
- ^ Weitz JI, Strony J, Ageno W, Gailani D, Hylek EM, Lassen MR, et al. (November 2021). "Milvexian for the Prevention of Venous Thromboembolism". The New England Journal of Medicine. 385 (23): 2161–2172. doi:10.1056/NEJMoa2113194. PMC 9540352. PMID 34780683. S2CID 244132392.
- ^ Long AT, Kenne E, Jung R, Fuchs TA, Renné T (March 2016). "Contact system revisited: an interface between inflammation, coagulation, and innate immunity". Journal of Thrombosis and Haemostasis. 14 (3): 427–437. doi:10.1111/jth.13235. PMID 26707513.
- ^ Verhamme P, Yi BA, Segers A, Salter J, Bloomfield D, Buller HR, et al. (12 August 2021). "Abelacimab for Prevention of Venous Thromboembolism". The New England Journal of Medicine. 385 (7): 609–617. doi:10.1056/NEJMoa2105872. PMID 34297496. S2CID 236198598.
- ^ Ruff CT, Patel SM, Giugliano RP, Morrow DA, Hug B, Kuder JF, et al. (23 January 2025). "Abelacimab versus Rivaroxaban in Patients with Atrial Fibrillation". The New England Journal of Medicine. 392 (4): 361–371. doi:10.1056/NEJMoa2406674. PMID 39842011.
Further reading
[edit]- Gailani D, Zivelin A, Sinha D, Walsh PN (2005). "Do platelets synthesize factor XI?". Journal of Thrombosis and Haemostasis. 2 (10): 1709–1712. doi:10.1111/j.1538-7836.2004.00935.x. PMID 15456479. S2CID 5962587.
- Dossenbach-Glaninger A, Hopmeier P (Jun 2005). "Coagulation factor XI: a database of mutations and polymorphisms associated with factor XI deficiency". Blood Coagulation & Fibrinolysis. 16 (4): 231–238. doi:10.1097/01.mbc.0000169214.62560.a5. PMID 15870541. S2CID 23922781.
- Seligsohn U (Jul 2007). "Factor XI in haemostasis and thrombosis: past, present and future". Thrombosis and Haemostasis. 98 (1): 84–89. doi:10.1160/th07-04-0246. PMID 17597996.
- Meijers JC, Davie EW, Chung DW (Mar 1992). "Expression of human blood coagulation factor XI: characterization of the defect in factor XI type III deficiency". Blood. 79 (6): 1435–1440. doi:10.1182/blood.V79.6.1435.1435. PMID 1547342.
- Gailani D, Broze GJ (Aug 1991). "Factor XI activation in a revised model of blood coagulation". Science. 253 (5022). New York, N.Y.: 909–912. Bibcode:1991Sci...253..909G. doi:10.1126/science.1652157. PMID 1652157. S2CID 9262836.
- Buetow KH, Shiang R, Yang P, Nakamura Y, Lathrop GM, White R, et al. (May 1991). "A detailed multipoint map of human chromosome 4 provides evidence for linkage heterogeneity and position-specific recombination rates". American Journal of Human Genetics. 48 (5): 911–925. PMC 1683054. PMID 1673289.
- Bodfish P, Warne D, Watkins C, Nyberg K, Spurr NK (1992). "Dinucleotide repeat polymorphism in the human coagulation factor XI gene, intron B (F11), detected using the polymerase chain reaction". Nucleic Acids Research. 19 (24): 6979. doi:10.1093/nar/19.24.6979-a. PMC 329377. PMID 1762944.
- Clarkson K, Rosenfeld B, Fair J, Klein A, Bell W (Dec 1991). "Factor XI deficiency acquired by liver transplantation". Annals of Internal Medicine. 115 (11): 877–879. doi:10.7326/0003-4819-115-11-877. PMID 1952475.
- McMullen BA, Fujikawa K, Davie EW (Feb 1991). "Location of the disulfide bonds in human coagulation factor XI: the presence of tandem apple domains". Biochemistry. 30 (8): 2056–2060. doi:10.1021/bi00222a008. PMID 1998667.
- Naito K, Fujikawa K (Apr 1991). "Activation of human blood coagulation factor XI independent of factor XII. Factor XI is activated by thrombin and factor XIa in the presence of negatively charged surfaces". The Journal of Biological Chemistry. 266 (12): 7353–7358. doi:10.1016/S0021-9258(20)89453-8. PMID 2019570.
- Asakai R, Chung DW, Davie EW, Seligsohn U (Jul 1991). "Factor XI deficiency in Ashkenazi Jews in Israel". The New England Journal of Medicine. 325 (3): 153–158. doi:10.1056/NEJM199107183250303. PMID 2052060.
- España F, Berrettini M, Griffin JH (Aug 1989). "Purification and characterization of plasma protein C inhibitor". Thrombosis Research. 55 (3): 369–384. doi:10.1016/0049-3848(89)90069-8. PMID 2551064.
- Asakai R, Chung DW, Ratnoff OD, Davie EW (Oct 1989). "Factor XI (plasma thromboplastin antecedent) deficiency in Ashkenazi Jews is a bleeding disorder that can result from three types of point mutations". Proceedings of the National Academy of Sciences of the United States of America. 86 (20): 7667–7671. Bibcode:1989PNAS...86.7667A. doi:10.1073/pnas.86.20.7667. PMC 298131. PMID 2813350.
- Asakai R, Davie EW, Chung DW (1988). "Organization of the gene for human factor XI". Biochemistry. 26 (23): 7221–7228. doi:10.1021/bi00397a004. PMID 2827746.
- Fujikawa K, Chung DW, Hendrickson LE, Davie EW (May 1986). "Amino acid sequence of human factor XI, a blood coagulation factor with four tandem repeats that are highly homologous with plasma prekallikrein". Biochemistry. 25 (9): 2417–2424. doi:10.1021/bi00357a018. PMID 3636155.
- Warn-Cramer BJ, Bajaj SP (1986). "Stoichiometry of binding of high molecular weight kininogen to factor XI/XIa". Biochemical and Biophysical Research Communications. 133 (2): 417–422. doi:10.1016/0006-291X(85)90922-2. PMID 3936495.
- Bouma BN, Vlooswijk RA, Griffin JH (Nov 1983). "Immunologic studies of human coagulation factor XI and its complex with high molecular weight kininogen". Blood. 62 (5): 1123–1131. doi:10.1182/blood.V62.5.1123.1123. PMID 6626744.
- Tuszynski GP, Bevacqua SJ, Schmaier AH, Colman RW, Walsh PN (Jun 1982). "Factor XI antigen and activity in human platelets". Blood. 59 (6): 1148–1156. doi:10.1182/blood.V59.6.1148.1148. PMID 7044446.
- Imanaka Y, Lal K, Nishimura T, Bolton-Maggs PH, Tuddenham EG, McVey JH (Aug 1995). "Identification of two novel mutations in non-Jewish factor XI deficiency". British Journal of Haematology. 90 (4): 916–920. doi:10.1111/j.1365-2141.1995.tb05215.x. PMID 7669672. S2CID 21900907.
- Pugh RE, McVey JH, Tuddenham EG, Hancock JF (Mar 1995). "Six point mutations that cause factor XI deficiency". Blood. 85 (6): 1509–1516. doi:10.1182/blood.V85.6.1509.bloodjournal8561509. PMID 7888672.
- Riley PW, Cheng H, Samuel D, Roder H, Walsh PN (Mar 2007). "Dimer dissociation and unfolding mechanism of coagulation factor XI apple 4 domain: spectroscopic and mutational analysis". Journal of Molecular Biology. 367 (2): 558–573. doi:10.1016/j.jmb.2006.12.066. PMC 1945241. PMID 17257616.
- Samuel D, Cheng H, Riley PW, Canutescu AA, Nagaswami C, Weisel JW, et al. (Oct 2007). "Solution structure of the A4 domain of factor XI sheds light on the mechanism of zymogen activation". Proceedings of the National Academy of Sciences of the United States of America. 104 (40): 15693–15698. Bibcode:2007PNAS..10415693S. doi:10.1073/pnas.0703080104. PMC 1987390. PMID 17884987.
External links
[edit]- The MEROPS online database for peptidases and their inhibitors: S01.213 Archived 2020-05-29 at the Wayback Machine
PDB gallery | |
|---|---|
|












![1zpb: Crystal Structure of the Catalytic Domain of Coagulation Factor XI in Complex with 4-Methyl-pentanoic acid {1-[4-guanidino-1-(thiazole-2-carbonyl)-butylcarbamoyl]-2-methyl-propyl}-amide](https://en.wikipedia.org/wiki/File:PDB_1zpb_EBI.png)
![1zpc: Crystal Structure of the Catalytic Domain of Coagulation Factor XI in Complex with 2-[2-(3-Chloro-phenyl)-2-hydroxy-acetylamino]-N-[4-guanidino-1-(thiazole-2-carbonyl)-butyl]-3-methyl-butyramide](https://en.wikipedia.org/wiki/File:PDB_1zpc_EBI.png)





![1ztj: Crystal Structure of the Catalytic Domain of Coagulation Factor XI in Complex with 2-(5-Benzylamino-2-methylsulfanyl-6-oxo-6H-pyrimidin-1-yl)-N-[4-guanidino-1-(thiazole-2-carbonyl)-butyl]-acetamide](https://en.wikipedia.org/wiki/File:PDB_1ztj_EBI.png)
![1ztk: Crystal Structure of the Catalytic Domain of Coagulation Factor XI in Complex with 2-(5-Amino-6-oxo-2-m-tolyl-6H-pyrimidin-1-yl)-N-[4-guanidino-1-(thiazole-2-carbonyl)-butyl]-acetamide](https://en.wikipedia.org/wiki/File:PDB_1ztk_EBI.png)
![1ztl: Crystal Structure of the Catalytic Domain of Coagulation Factor XI in Complex with N-[4-Guanidino-1-(thiazole-2-carbonyl)-butyl]-2-{6-oxo-5-[(quinolin-8-ylmethyl)-amino]-2-m-tolyl-6H-pyrimidin-1-yl}-acetamide](https://en.wikipedia.org/wiki/File:PDB_1ztl_EBI.png)

