Recent from talks
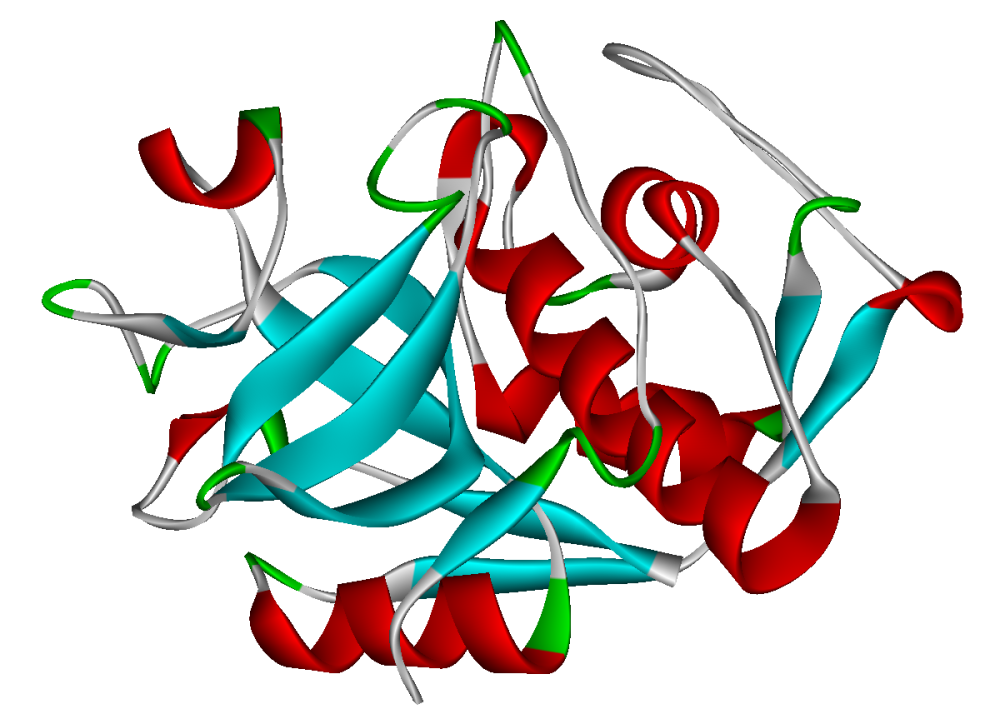
Cathepsin
Knowledge base stats:
Talk channels stats:
Members stats:
Cathepsin
Cathepsins (Ancient Greek kata- "down" and hepsein "boil"; abbreviated CTS) are proteases (enzymes that degrade proteins) found in all animals as well as other organisms. There are approximately a dozen members of this family, which are distinguished by their structure, catalytic mechanism, and which proteins they cleave[citation needed]. Most of the members become activated at the low pH found in lysosomes. Thus, the activity of this family lies almost entirely within those organelles. There are, however, exceptions such as cathepsin K, which works extracellularly after secretion by osteoclasts in bone resorption. Cathepsins have a vital role in mammalian cellular turnover.
Cathepsins are involved in many physiological processes and have been implicated in a number of human diseases. The cysteine cathepsins have attracted significant research effort as drug targets.
Deficiencies in this protein are linked to multiple forms of galactosialidosis. The cathepsin A activity in lysates of metastatic lesions of malignant melanoma is significantly higher than in primary focus lysates. Cathepsin A increased in muscles moderately affected by muscular dystrophy and denervating diseases.
Cathepsin B may function as a beta-secretase 1, cleaving amyloid precursor protein to produce amyloid beta. Overexpression of the encoded protein, which is a member of the peptidase C1 family, has been associated with esophageal adenocarcinoma and other tumors. Cathepsin B has also been implicated in the progression of various human tumors including ovarian cancer.
Cathepsin D (an aspartyl protease) appears to cleave a variety of substrates such as fibronectin and laminin. Unlike some of the other cathepsins, cathepsin D has some protease activity at neutral pH. High levels of this enzyme in tumor cells seems to be associated with greater invasiveness.
Cathepsin K is the most potent mammalian collagenase. Cathepsin K is involved in osteoporosis, a disease in which a decrease in bone density causes an increased risk for fracture. Osteoclasts are the bone resorbing cells of the body, and they secrete cathepsin K in order to break down collagen, the major component of the non-mineral protein matrix of the bone. Cathepsin K, among other cathepsins, plays a role in cancer metastasis through the degradation of the extracellular matrix. The genetic knockout for cathepsin S and K in mice with atherosclerosis was shown to reduce the size of atherosclerotic lesions. The expression of cathepsin K in cultured endothelial cells is regulated by shear stress. Cathepsin K has also been shown to play a role in arthritis.
Mouse cathepsin L is homologous to human cathepsin V. Mouse cathepsin L has been shown to play a role in adipogenesis and glucose intolerance in mice. Cathepsin L degrades fibronectin, insulin receptor (IR), and insulin-like growth factor 1 receptor (IGF-1R). Cathepsin L-deficient mice were shown to have less adipose tissue, lower serum glucose and insulin levels, more insulin receptor subunits, more glucose transporter (GLUT4) and more fibronectin than wild type controls.
Five cyclic peptides show inhibitory activity towards human cathepsins L, B, H, and K. Several inhibitors have reached clinical trials, targeting cathepsins K and S as promising therapeutics for osteoporosis, osteoarthritis, and chronic pain. Cathepsin K inhibitors, Relacatib, Balicatib, and Odanacatib, were terminated during clinical trials at phases I, II, and III, respectively, owing to adverse side effects. SAR114137, a Cathepsin S inhibitor, did not progress past phase I for chronic pain. In 2022, STI-1558, a Cathepsin L inhibitor, received FDA clearance to begin phase I studies to treat COVID-19.
Hub AI
Cathepsin AI simulator
(@Cathepsin_simulator)
Cathepsin
Cathepsins (Ancient Greek kata- "down" and hepsein "boil"; abbreviated CTS) are proteases (enzymes that degrade proteins) found in all animals as well as other organisms. There are approximately a dozen members of this family, which are distinguished by their structure, catalytic mechanism, and which proteins they cleave[citation needed]. Most of the members become activated at the low pH found in lysosomes. Thus, the activity of this family lies almost entirely within those organelles. There are, however, exceptions such as cathepsin K, which works extracellularly after secretion by osteoclasts in bone resorption. Cathepsins have a vital role in mammalian cellular turnover.
Cathepsins are involved in many physiological processes and have been implicated in a number of human diseases. The cysteine cathepsins have attracted significant research effort as drug targets.
Deficiencies in this protein are linked to multiple forms of galactosialidosis. The cathepsin A activity in lysates of metastatic lesions of malignant melanoma is significantly higher than in primary focus lysates. Cathepsin A increased in muscles moderately affected by muscular dystrophy and denervating diseases.
Cathepsin B may function as a beta-secretase 1, cleaving amyloid precursor protein to produce amyloid beta. Overexpression of the encoded protein, which is a member of the peptidase C1 family, has been associated with esophageal adenocarcinoma and other tumors. Cathepsin B has also been implicated in the progression of various human tumors including ovarian cancer.
Cathepsin D (an aspartyl protease) appears to cleave a variety of substrates such as fibronectin and laminin. Unlike some of the other cathepsins, cathepsin D has some protease activity at neutral pH. High levels of this enzyme in tumor cells seems to be associated with greater invasiveness.
Cathepsin K is the most potent mammalian collagenase. Cathepsin K is involved in osteoporosis, a disease in which a decrease in bone density causes an increased risk for fracture. Osteoclasts are the bone resorbing cells of the body, and they secrete cathepsin K in order to break down collagen, the major component of the non-mineral protein matrix of the bone. Cathepsin K, among other cathepsins, plays a role in cancer metastasis through the degradation of the extracellular matrix. The genetic knockout for cathepsin S and K in mice with atherosclerosis was shown to reduce the size of atherosclerotic lesions. The expression of cathepsin K in cultured endothelial cells is regulated by shear stress. Cathepsin K has also been shown to play a role in arthritis.
Mouse cathepsin L is homologous to human cathepsin V. Mouse cathepsin L has been shown to play a role in adipogenesis and glucose intolerance in mice. Cathepsin L degrades fibronectin, insulin receptor (IR), and insulin-like growth factor 1 receptor (IGF-1R). Cathepsin L-deficient mice were shown to have less adipose tissue, lower serum glucose and insulin levels, more insulin receptor subunits, more glucose transporter (GLUT4) and more fibronectin than wild type controls.
Five cyclic peptides show inhibitory activity towards human cathepsins L, B, H, and K. Several inhibitors have reached clinical trials, targeting cathepsins K and S as promising therapeutics for osteoporosis, osteoarthritis, and chronic pain. Cathepsin K inhibitors, Relacatib, Balicatib, and Odanacatib, were terminated during clinical trials at phases I, II, and III, respectively, owing to adverse side effects. SAR114137, a Cathepsin S inhibitor, did not progress past phase I for chronic pain. In 2022, STI-1558, a Cathepsin L inhibitor, received FDA clearance to begin phase I studies to treat COVID-19.
Recent media