Recent from talks
Retina horizontal cell
Knowledge base stats:
Talk channels stats:
Members stats:
Retina horizontal cell
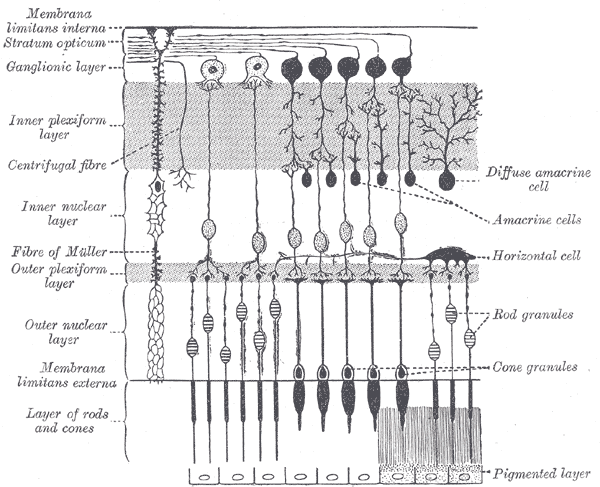
Horizontal cells are the laterally interconnecting neurons having cell bodies in the inner nuclear layer of the retina of vertebrate eyes. They help integrate and regulate the input from multiple photoreceptor cells. Among their functions, horizontal cells are believed to be responsible for increasing contrast via lateral inhibition and adapting both to bright and dim light conditions. Horizontal cells provide inhibitory feedback to rod and cone photoreceptors. They are thought to be important for the antagonistic center-surround property of the receptive fields of many types of retinal ganglion cells.
Other retinal neurons include photoreceptor cells, bipolar cells, amacrine cells, and retinal ganglion cells.
Depending on the species, there are typically one or two classes of horizontal cells, with a third type sometimes proposed.
Horizontal cells span across photoreceptors and summate inputs before synapsing onto photoreceptor cells. Horizontal cells may also synapse onto bipolar cells, but this remains uncertain.
There is a greater density of horizontal cells towards the central region of the retina. In the cat, it is observed that A-type horizontal cells have a density of 225 cells/mm2 near the center of the retina and a density of 120 cells/mm2 in more peripheral retina.
Horizontal cells and other retinal interneuron cells are less likely to be near neighbours of the same subtype than would occur by chance, resulting in 'exclusion zones' that separate them. Mosaic arrangements provide a mechanism to distribute each cell type evenly across the retina, ensuring that all parts of the visual field have access to a full set of processing elements. MEGF10 and MEGF11 transmembrane proteins have critical roles in the formation of the mosaics by horizontal cells and starburst amacrine cells in mice.
Horizontal cells are depolarized by the release of glutamate from photoreceptors, which happens in the absence of light. Depolarization of a horizontal cell causes it to hyperpolarize nearby photoreceptors. Conversely, in the light, a photoreceptor releases less glutamate, which hyperpolarizes the horizontal cell, leading to depolarization of nearby photoreceptors. Thus, horizontal cells provide negative feedback to photoreceptors. The moderately wide lateral spread and coupling of horizontal cells by gap junctions, measures the average level of illumination falling upon a region of the retinal surface, which horizontal cells then subtract a proportionate value from the output of photoreceptors to hold the signal input to the inner retinal circuitry within its operating range. Horizontal cells are also one of two groups of inhibitory interneurons that contribute to the surround of retinal ganglion cells:
Illumination Center photoreceptor hyperpolarization Horizontal cell hyperpolarization Surround photoreceptor depolarization

Hub AI
Retina horizontal cell AI simulator
(@Retina horizontal cell_simulator)
Retina horizontal cell
Horizontal cells are the laterally interconnecting neurons having cell bodies in the inner nuclear layer of the retina of vertebrate eyes. They help integrate and regulate the input from multiple photoreceptor cells. Among their functions, horizontal cells are believed to be responsible for increasing contrast via lateral inhibition and adapting both to bright and dim light conditions. Horizontal cells provide inhibitory feedback to rod and cone photoreceptors. They are thought to be important for the antagonistic center-surround property of the receptive fields of many types of retinal ganglion cells.
Other retinal neurons include photoreceptor cells, bipolar cells, amacrine cells, and retinal ganglion cells.
Depending on the species, there are typically one or two classes of horizontal cells, with a third type sometimes proposed.
Horizontal cells span across photoreceptors and summate inputs before synapsing onto photoreceptor cells. Horizontal cells may also synapse onto bipolar cells, but this remains uncertain.
There is a greater density of horizontal cells towards the central region of the retina. In the cat, it is observed that A-type horizontal cells have a density of 225 cells/mm2 near the center of the retina and a density of 120 cells/mm2 in more peripheral retina.
Horizontal cells and other retinal interneuron cells are less likely to be near neighbours of the same subtype than would occur by chance, resulting in 'exclusion zones' that separate them. Mosaic arrangements provide a mechanism to distribute each cell type evenly across the retina, ensuring that all parts of the visual field have access to a full set of processing elements. MEGF10 and MEGF11 transmembrane proteins have critical roles in the formation of the mosaics by horizontal cells and starburst amacrine cells in mice.
Horizontal cells are depolarized by the release of glutamate from photoreceptors, which happens in the absence of light. Depolarization of a horizontal cell causes it to hyperpolarize nearby photoreceptors. Conversely, in the light, a photoreceptor releases less glutamate, which hyperpolarizes the horizontal cell, leading to depolarization of nearby photoreceptors. Thus, horizontal cells provide negative feedback to photoreceptors. The moderately wide lateral spread and coupling of horizontal cells by gap junctions, measures the average level of illumination falling upon a region of the retinal surface, which horizontal cells then subtract a proportionate value from the output of photoreceptors to hold the signal input to the inner retinal circuitry within its operating range. Horizontal cells are also one of two groups of inhibitory interneurons that contribute to the surround of retinal ganglion cells:
Illumination Center photoreceptor hyperpolarization Horizontal cell hyperpolarization Surround photoreceptor depolarization
Recent media