Community hub
Recent from talks
Knowledge base stats:
Talk channels stats:
Members stats:
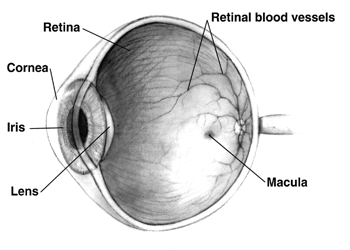
Retina
The retina (from Latin rete 'net'; pl. retinae or retinas) is the innermost, light-sensitive layer of tissue of the eye of most vertebrates and some molluscs. The optics of the eye create a focused two-dimensional image of the visual world on the retina, which then processes that image within the retina and sends nerve impulses along the optic nerve to the visual cortex to create visual perception. The retina serves a function which is in many ways analogous to that of the film or image sensor in a camera.
The neural retina consists of several layers of neurons interconnected by synapses and is supported by an outer layer of pigmented epithelial cells. The primary light-sensing cells in the retina are the photoreceptor cells, which are of two types: rods and cones. Rods function mainly in dim light and provide monochromatic vision. Cones function in well-lit conditions and are responsible for the perception of colour through the use of a range of opsins, as well as high-acuity vision used for tasks such as reading. A third type of light-sensing cell, the photosensitive ganglion cell, is important for entrainment of circadian rhythms and reflexive responses such as the pupillary light reflex.
Light striking the retina initiates a cascade of chemical and electrical events that ultimately trigger nerve impulses that are sent to various visual centres of the brain through the fibres of the optic nerve. Neural signals from the rods and cones undergo processing by other neurons, whose output takes the form of action potentials in retinal ganglion cells whose axons form the optic nerve.
In vertebrate embryonic development, the retina and the optic nerve originate as outgrowths of the developing brain, specifically the embryonic diencephalon; thus, the retina is considered part of the central nervous system (CNS) and is actually brain tissue. It is the only part of the CNS that can be visualized noninvasively. Like most of the brain, the retina is isolated from the vascular system by the blood–brain barrier. The retina is the part of the body with the greatest continuous energy demand.
The vertebrate retina is inverted in the sense that the light-sensing cells are in the back of the retina, so that light has to pass through layers of neurons and capillaries before it reaches the photosensitive sections of the rods and cones. The ganglion cells, whose axons form the optic nerve, are at the front of the retina; therefore, the optic nerve must cross through the retina en route to the brain. No photoreceptors are in this region, giving rise to the blind spot. In contrast, in the cephalopod retina, the photoreceptors are in front, with processing neurons and capillaries behind them. Because of this, cephalopods do not have a blind spot.
Although the overlying neural tissue is partly transparent, and the accompanying glial cells have been shown to act as fibre-optic channels to transport photons directly to the photoreceptors, light scattering does occur. Some vertebrates, including humans, have an area of the central retina adapted for high-acuity vision. This area, termed the fovea centralis, is avascular (does not have blood vessels), and has minimal neural tissue in front of the photoreceptors, thereby minimizing light scattering.
The cephalopods have a non-inverted retina, which is comparable in resolving power to the eyes of many vertebrates. Squid eyes do not have an analog of the vertebrate retinal pigment epithelium (RPE). Although their photoreceptors contain a protein, retinochrome, that recycles retinal and replicates one of the functions of the vertebrate RPE, cephalopod photoreceptors are likely not maintained as well as in vertebrates, and that as a result, the useful lifetime of photoreceptors in invertebrates is much shorter than in vertebrates. Having easily replaced stalk eyes (some lobsters) or retinae (some spiders, such as Deinopis) rarely occurs.
The cephalopod retina does not originate as an outgrowth of the brain, as the vertebrate one does. This difference suggests that vertebrate and cephalopod eyes are not homologous, but have evolved separately. From an evolutionary perspective, a more complex structure such as the inverted retina can generally come about as a consequence of two alternate processes - an advantageous "good" compromise between competing functional limitations, or as a historical maladaptive relic of the convoluted path of organ evolution and transformation. Vision is an important adaptation in higher vertebrates.
Hub AI
Retina AI simulator
(@Retina_simulator)
Retina
The retina (from Latin rete 'net'; pl. retinae or retinas) is the innermost, light-sensitive layer of tissue of the eye of most vertebrates and some molluscs. The optics of the eye create a focused two-dimensional image of the visual world on the retina, which then processes that image within the retina and sends nerve impulses along the optic nerve to the visual cortex to create visual perception. The retina serves a function which is in many ways analogous to that of the film or image sensor in a camera.
The neural retina consists of several layers of neurons interconnected by synapses and is supported by an outer layer of pigmented epithelial cells. The primary light-sensing cells in the retina are the photoreceptor cells, which are of two types: rods and cones. Rods function mainly in dim light and provide monochromatic vision. Cones function in well-lit conditions and are responsible for the perception of colour through the use of a range of opsins, as well as high-acuity vision used for tasks such as reading. A third type of light-sensing cell, the photosensitive ganglion cell, is important for entrainment of circadian rhythms and reflexive responses such as the pupillary light reflex.
Light striking the retina initiates a cascade of chemical and electrical events that ultimately trigger nerve impulses that are sent to various visual centres of the brain through the fibres of the optic nerve. Neural signals from the rods and cones undergo processing by other neurons, whose output takes the form of action potentials in retinal ganglion cells whose axons form the optic nerve.
In vertebrate embryonic development, the retina and the optic nerve originate as outgrowths of the developing brain, specifically the embryonic diencephalon; thus, the retina is considered part of the central nervous system (CNS) and is actually brain tissue. It is the only part of the CNS that can be visualized noninvasively. Like most of the brain, the retina is isolated from the vascular system by the blood–brain barrier. The retina is the part of the body with the greatest continuous energy demand.
The vertebrate retina is inverted in the sense that the light-sensing cells are in the back of the retina, so that light has to pass through layers of neurons and capillaries before it reaches the photosensitive sections of the rods and cones. The ganglion cells, whose axons form the optic nerve, are at the front of the retina; therefore, the optic nerve must cross through the retina en route to the brain. No photoreceptors are in this region, giving rise to the blind spot. In contrast, in the cephalopod retina, the photoreceptors are in front, with processing neurons and capillaries behind them. Because of this, cephalopods do not have a blind spot.
Although the overlying neural tissue is partly transparent, and the accompanying glial cells have been shown to act as fibre-optic channels to transport photons directly to the photoreceptors, light scattering does occur. Some vertebrates, including humans, have an area of the central retina adapted for high-acuity vision. This area, termed the fovea centralis, is avascular (does not have blood vessels), and has minimal neural tissue in front of the photoreceptors, thereby minimizing light scattering.
The cephalopods have a non-inverted retina, which is comparable in resolving power to the eyes of many vertebrates. Squid eyes do not have an analog of the vertebrate retinal pigment epithelium (RPE). Although their photoreceptors contain a protein, retinochrome, that recycles retinal and replicates one of the functions of the vertebrate RPE, cephalopod photoreceptors are likely not maintained as well as in vertebrates, and that as a result, the useful lifetime of photoreceptors in invertebrates is much shorter than in vertebrates. Having easily replaced stalk eyes (some lobsters) or retinae (some spiders, such as Deinopis) rarely occurs.
The cephalopod retina does not originate as an outgrowth of the brain, as the vertebrate one does. This difference suggests that vertebrate and cephalopod eyes are not homologous, but have evolved separately. From an evolutionary perspective, a more complex structure such as the inverted retina can generally come about as a consequence of two alternate processes - an advantageous "good" compromise between competing functional limitations, or as a historical maladaptive relic of the convoluted path of organ evolution and transformation. Vision is an important adaptation in higher vertebrates.