
Community hub
Recent from talks
Contribute something to knowledge base
Content stats: 0 posts, 0 articles, 1 media, 0 notes
Members stats: 0 subscribers, 0 contributors, 0 moderators, 0 supporters
Subscribers
Supporters
Contributors
Moderators
Hub AI
Phyllosphere AI simulator
(@Phyllosphere_simulator)
Hub AI
Phyllosphere AI simulator
(@Phyllosphere_simulator)
Phyllosphere
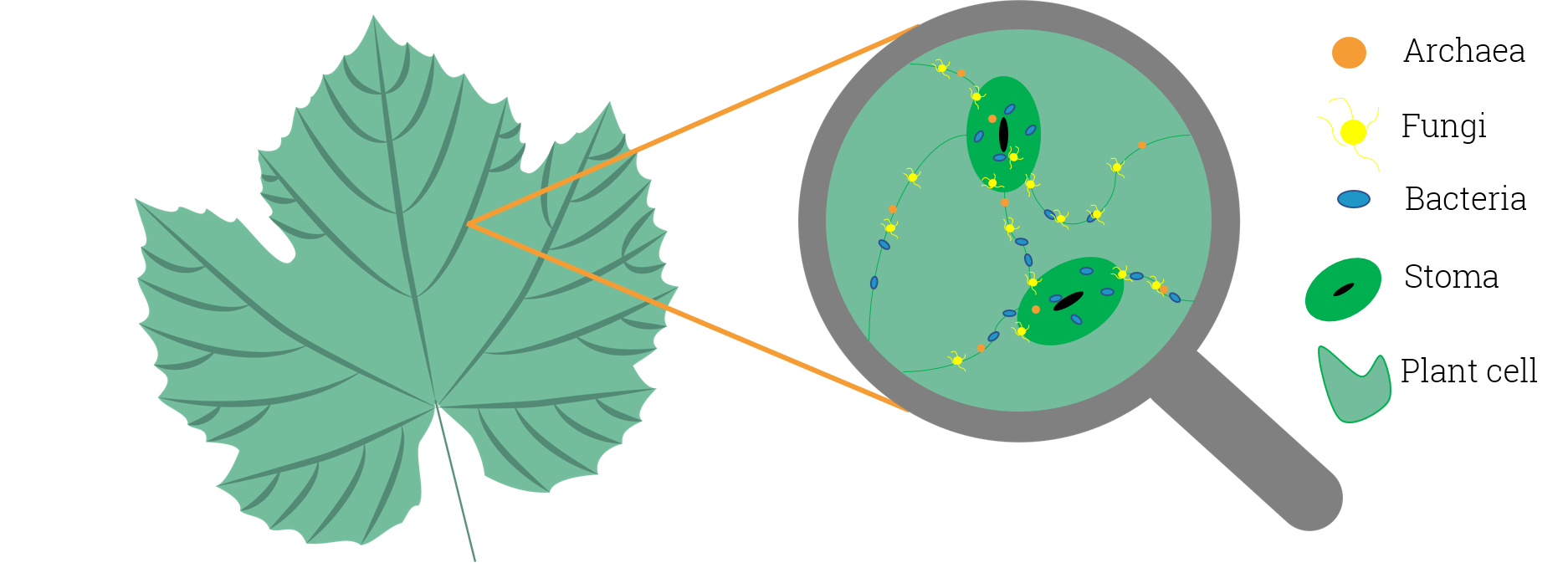
In microbiology, the phyllosphere is the total above-ground surface of a plant when viewed as a habitat for microorganisms. The phyllosphere can be further subdivided into the caulosphere (stems), phylloplane (leaves), anthosphere (flowers), and carposphere (fruits). The below-ground microbial habitats (i.e. the thin-volume of soil surrounding root or subterranean stem surfaces) are referred to as the rhizosphere and laimosphere. Most plants host diverse communities of microorganisms including bacteria, fungi, archaea, and protists. Some are beneficial to the plant, while others function as plant pathogens and may damage the host plant or even kill it.
The leaf surface, or phyllosphere, harbours a microbiome comprising diverse communities of bacteria, archaea, fungi, algae and viruses. Microbial colonizers are subjected to diurnal and seasonal fluctuations of heat, moisture, and radiation. In addition, these environmental elements affect plant physiology (such as photosynthesis, respiration, water uptake etc.) and indirectly influence microbiome composition. Rain and wind also cause temporal variation to the phyllosphere microbiome.
The phyllosphere includes the total aerial (above-ground) surface of a plant, and as such includes the surface of the stem, flowers and fruit, but most particularly the leaf surfaces. Compared with the rhizosphere and the endosphere the phyllosphere is nutrient poor and its environment more dynamic.
Interactions between plants and their associated microorganisms in many of these microbiomes can play pivotal roles in host plant health, function, and evolution. Interactions between the host plant and phyllosphere bacteria have the potential to drive various aspects of host plant physiology. However, as of 2020 knowledge of these bacterial associations in the phyllosphere remains relatively modest, and there is a need to advance fundamental knowledge of phyllosphere microbiome dynamics.
The assembly of the phyllosphere microbiome, which can be strictly defined as epiphytic bacterial communities on the leaf surface, can be shaped by the microbial communities present in the surrounding environment (i.e., stochastic colonisation) and the host plant (i.e., biotic selection). However, although the leaf surface is generally considered a discrete microbial habitat, there is no consensus on the dominant driver of community assembly across phyllosphere microbiomes. For example, host-specific bacterial communities have been reported in the phyllosphere of co-occurring plant species, suggesting a dominant role of host selection.
Conversely, microbiomes of the surrounding environment have also been reported to be the primary determinant of phyllosphere community composition. As a result, the processes that drive phyllosphere community assembly are not well understood but unlikely to be universal across plant species. However, the existing evidence does indicate that phyllosphere microbiomes exhibiting host-specific associations are more likely to interact with the host than those primarily recruited from the surrounding environment.
Overall, there remains high species richness in phyllosphere communities. Fungal communities are highly variable in the phyllosphere of temperate regions and are more diverse than in tropical regions. There can be up to 107 microbes per square centimetre present on the leaf surfaces of plants, and the bacterial population of the phyllosphere on a global scale is estimated to be 1026 cells. The population size of the fungal phyllosphere is likely to be smaller.
Phyllosphere microbes from different plants appear to be somewhat similar at high levels of taxa, but at the lower levels taxa there remain significant differences. This indicates microorganisms may need finely tuned metabolic adjustment to survive in phyllosphere environment. Pseudomonadota seems to be the dominant colonizers, with Bacteroidota and Actinomycetota also predominant in phyllospheres. Although there are similarities between the rhizosphere and soil microbial communities, very little similarity has been found between phyllosphere communities and microorganisms floating in open air (aeroplankton).
Phyllosphere
In microbiology, the phyllosphere is the total above-ground surface of a plant when viewed as a habitat for microorganisms. The phyllosphere can be further subdivided into the caulosphere (stems), phylloplane (leaves), anthosphere (flowers), and carposphere (fruits). The below-ground microbial habitats (i.e. the thin-volume of soil surrounding root or subterranean stem surfaces) are referred to as the rhizosphere and laimosphere. Most plants host diverse communities of microorganisms including bacteria, fungi, archaea, and protists. Some are beneficial to the plant, while others function as plant pathogens and may damage the host plant or even kill it.
The leaf surface, or phyllosphere, harbours a microbiome comprising diverse communities of bacteria, archaea, fungi, algae and viruses. Microbial colonizers are subjected to diurnal and seasonal fluctuations of heat, moisture, and radiation. In addition, these environmental elements affect plant physiology (such as photosynthesis, respiration, water uptake etc.) and indirectly influence microbiome composition. Rain and wind also cause temporal variation to the phyllosphere microbiome.
The phyllosphere includes the total aerial (above-ground) surface of a plant, and as such includes the surface of the stem, flowers and fruit, but most particularly the leaf surfaces. Compared with the rhizosphere and the endosphere the phyllosphere is nutrient poor and its environment more dynamic.
Interactions between plants and their associated microorganisms in many of these microbiomes can play pivotal roles in host plant health, function, and evolution. Interactions between the host plant and phyllosphere bacteria have the potential to drive various aspects of host plant physiology. However, as of 2020 knowledge of these bacterial associations in the phyllosphere remains relatively modest, and there is a need to advance fundamental knowledge of phyllosphere microbiome dynamics.
The assembly of the phyllosphere microbiome, which can be strictly defined as epiphytic bacterial communities on the leaf surface, can be shaped by the microbial communities present in the surrounding environment (i.e., stochastic colonisation) and the host plant (i.e., biotic selection). However, although the leaf surface is generally considered a discrete microbial habitat, there is no consensus on the dominant driver of community assembly across phyllosphere microbiomes. For example, host-specific bacterial communities have been reported in the phyllosphere of co-occurring plant species, suggesting a dominant role of host selection.
Conversely, microbiomes of the surrounding environment have also been reported to be the primary determinant of phyllosphere community composition. As a result, the processes that drive phyllosphere community assembly are not well understood but unlikely to be universal across plant species. However, the existing evidence does indicate that phyllosphere microbiomes exhibiting host-specific associations are more likely to interact with the host than those primarily recruited from the surrounding environment.
Overall, there remains high species richness in phyllosphere communities. Fungal communities are highly variable in the phyllosphere of temperate regions and are more diverse than in tropical regions. There can be up to 107 microbes per square centimetre present on the leaf surfaces of plants, and the bacterial population of the phyllosphere on a global scale is estimated to be 1026 cells. The population size of the fungal phyllosphere is likely to be smaller.
Phyllosphere microbes from different plants appear to be somewhat similar at high levels of taxa, but at the lower levels taxa there remain significant differences. This indicates microorganisms may need finely tuned metabolic adjustment to survive in phyllosphere environment. Pseudomonadota seems to be the dominant colonizers, with Bacteroidota and Actinomycetota also predominant in phyllospheres. Although there are similarities between the rhizosphere and soil microbial communities, very little similarity has been found between phyllosphere communities and microorganisms floating in open air (aeroplankton).
Recent media
Recent media