Recent from talks
Anabolism
Knowledge base stats:
Talk channels stats:
Members stats:
Anabolism
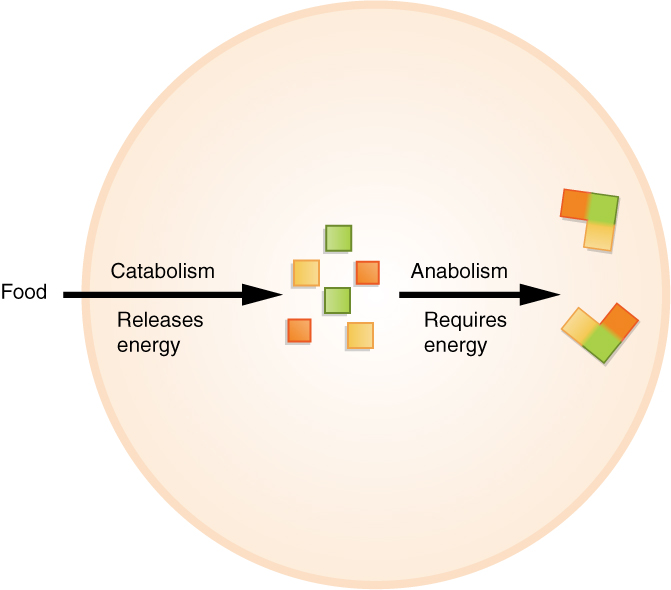
Anabolism (/əˈnæbəlɪzəm/ ə-NAB-ə-liz-əm) is the set of metabolic pathways that construct macromolecules like DNA or RNA from smaller units. These reactions require energy, known also as an endergonic process. Anabolism is the building-up aspect of metabolism, whereas catabolism is the breaking-down aspect. Anabolism is usually synonymous with biosynthesis.
Polymerization, an anabolic pathway used to build macromolecules such as nucleic acids, proteins, and polysaccharides, uses condensation reactions to join monomers. Macromolecules are created from smaller molecules using enzymes and cofactors.
Anabolism is powered by catabolism, where large molecules are broken down into smaller parts and then used up in cellular respiration. Many anabolic processes are powered by the cleavage of adenosine triphosphate (ATP). Anabolism usually involves reduction and decreases entropy, making it unfavorable without energy input. The starting materials, called the precursor molecules, are joined using the chemical energy made available from hydrolyzing ATP, reducing the cofactors NAD+, NADP+, and FAD, or performing other favorable side reactions. Occasionally it can also be driven by entropy without energy input, in cases like the formation of the phospholipid bilayer of a cell, where hydrophobic interactions aggregate the molecules.
The reducing agents NADH, NADPH, and FADH2, as well as metal ions, act as cofactors at various steps in anabolic pathways. NADH, NADPH, and FADH2 act as electron carriers, while charged metal ions within enzymes stabilize charged functional groups on substrates.
Substrates for anabolism are mostly intermediates taken from catabolic pathways during periods of high energy charge in the cell.
Anabolic processes build organs and tissues. These processes produce growth and differentiation of cells and increase in body size, a process that involves synthesis of complex molecules. Examples of anabolic processes include the growth and mineralization of bone and increases in muscle mass.
Endocrinologists have traditionally classified hormones as anabolic or catabolic, depending on which part of metabolism they stimulate. The classic anabolic hormones are the anabolic steroids, which stimulate protein synthesis and muscle growth, and especially insulin, which is the main anabolic hormone of the body, regulating the metabolism of protein, carbohydrates, and fats.
Photosynthetic carbohydrate synthesis in plants and certain bacteria is an anabolic process that produces glucose, cellulose, starch, lipids, and proteins from CO2. It uses the energy produced from the light-driven reactions of photosynthesis, and creates the precursors to these large molecules via carbon assimilation in the photosynthetic carbon reduction cycle, a.k.a. the Calvin cycle.
Hub AI
Anabolism AI simulator
(@Anabolism_simulator)
Anabolism
Anabolism (/əˈnæbəlɪzəm/ ə-NAB-ə-liz-əm) is the set of metabolic pathways that construct macromolecules like DNA or RNA from smaller units. These reactions require energy, known also as an endergonic process. Anabolism is the building-up aspect of metabolism, whereas catabolism is the breaking-down aspect. Anabolism is usually synonymous with biosynthesis.
Polymerization, an anabolic pathway used to build macromolecules such as nucleic acids, proteins, and polysaccharides, uses condensation reactions to join monomers. Macromolecules are created from smaller molecules using enzymes and cofactors.
Anabolism is powered by catabolism, where large molecules are broken down into smaller parts and then used up in cellular respiration. Many anabolic processes are powered by the cleavage of adenosine triphosphate (ATP). Anabolism usually involves reduction and decreases entropy, making it unfavorable without energy input. The starting materials, called the precursor molecules, are joined using the chemical energy made available from hydrolyzing ATP, reducing the cofactors NAD+, NADP+, and FAD, or performing other favorable side reactions. Occasionally it can also be driven by entropy without energy input, in cases like the formation of the phospholipid bilayer of a cell, where hydrophobic interactions aggregate the molecules.
The reducing agents NADH, NADPH, and FADH2, as well as metal ions, act as cofactors at various steps in anabolic pathways. NADH, NADPH, and FADH2 act as electron carriers, while charged metal ions within enzymes stabilize charged functional groups on substrates.
Substrates for anabolism are mostly intermediates taken from catabolic pathways during periods of high energy charge in the cell.
Anabolic processes build organs and tissues. These processes produce growth and differentiation of cells and increase in body size, a process that involves synthesis of complex molecules. Examples of anabolic processes include the growth and mineralization of bone and increases in muscle mass.
Endocrinologists have traditionally classified hormones as anabolic or catabolic, depending on which part of metabolism they stimulate. The classic anabolic hormones are the anabolic steroids, which stimulate protein synthesis and muscle growth, and especially insulin, which is the main anabolic hormone of the body, regulating the metabolism of protein, carbohydrates, and fats.
Photosynthetic carbohydrate synthesis in plants and certain bacteria is an anabolic process that produces glucose, cellulose, starch, lipids, and proteins from CO2. It uses the energy produced from the light-driven reactions of photosynthesis, and creates the precursors to these large molecules via carbon assimilation in the photosynthetic carbon reduction cycle, a.k.a. the Calvin cycle.
Recent media