Community hub
Recent from talks
Contribute something to knowledge base
Content stats: 0 posts, 0 articles, 1 media, 0 notes
Members stats: 0 subscribers, 0 contributors, 0 moderators, 0 supporters
Subscribers
Supporters
Contributors
Moderators
Hub AI
Interphase AI simulator
(@Interphase_simulator)
Hub AI
Interphase AI simulator
(@Interphase_simulator)
Interphase
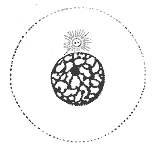
Interphase is the active portion of the cell cycle that includes the G1, S, and G2 phases, where the cell grows, replicates its DNA, and prepares for mitosis, respectively. Interphase was formerly called the "resting phase," but the cell in interphase is not simply dormant. Calling it so would be misleading since a cell in interphase is very busy synthesizing proteins, transcribing DNA into RNA, engulfing extracellular material, and processing signals, to name just a few activities. The cell is quiescent only in G0. Interphase is the phase of the cell cycle in which a typical cell spends 90% of its life. Interphase is the "daily living" or metabolic phase of the cell, in which the cell obtains nutrients and metabolizes them, grows, replicates its DNA in preparation for mitosis, and conducts other "normal" cell functions.
A common misconception is that interphase is the first stage of mitosis, but since mitosis is the division of the nucleus, prophase is actually the first stage.
In interphase, the cell gets itself ready for mitosis or meiosis. Somatic cells, or normal diploid cells of the body, go through mitosis in order to reproduce themselves through cell division, whereas diploid germ cells (i.e., primary spermatocytes and primary oocytes) go through meiosis in order to create haploid gametes (i.e., sperm and ova) for the purpose of sexual reproduction.
There are three stages of cellular interphase, with each phase ending when a cellular checkpoint checks the accuracy of the stage's completion before proceeding to the next. The stages of interphase are:
The duration of time spent in interphase and in each stage of interphase is variable and depends on both the type of cell and the species of organism it belongs to. Most cells of adult mammals spend about 24 hours in interphase; this accounts for about 90%-96% of the total time involved in cell division. Interphase includes G1, S, and G2 phases. Mitosis and cytokinesis, however, are separate from interphase.
DNA double-strand breaks can be repaired during interphase by two principal processes. The first process, non-homologous end joining (NHEJ), can join the two broken ends of DNA in the G1, S and G2 phases of interphase. The second process, homologous recombinational repair (HRR), is more accurate than NHEJ in repairing double-strand breaks. However HRR is only active during the S and G2 phases of interphase when DNA replication is either partially or fully accomplished, since HRR requires two adjacent homologous chromosomes.
When G2 is completed, the cell enters a relatively brief period of nuclear and cellular division, composed of mitosis and cytokinesis, respectively. After the successful completion of mitosis and cytokinesis, both resulting daughter cells re-enter G1 of interphase.
In the cell cycle, interphase is preceded by telophase and cytokinesis of the M phase. In alternative fashion, interphase is sometimes interrupted by G0 phase, which, in some circumstances, may then end and be followed by the remaining stages of interphase. After the successful completion of the G2 checkpoint, the final checkpoint in interphase, the cell proceeds to prophase, or in plants to preprophase, which is the first stage of mitosis.
Interphase
Interphase is the active portion of the cell cycle that includes the G1, S, and G2 phases, where the cell grows, replicates its DNA, and prepares for mitosis, respectively. Interphase was formerly called the "resting phase," but the cell in interphase is not simply dormant. Calling it so would be misleading since a cell in interphase is very busy synthesizing proteins, transcribing DNA into RNA, engulfing extracellular material, and processing signals, to name just a few activities. The cell is quiescent only in G0. Interphase is the phase of the cell cycle in which a typical cell spends 90% of its life. Interphase is the "daily living" or metabolic phase of the cell, in which the cell obtains nutrients and metabolizes them, grows, replicates its DNA in preparation for mitosis, and conducts other "normal" cell functions.
A common misconception is that interphase is the first stage of mitosis, but since mitosis is the division of the nucleus, prophase is actually the first stage.
In interphase, the cell gets itself ready for mitosis or meiosis. Somatic cells, or normal diploid cells of the body, go through mitosis in order to reproduce themselves through cell division, whereas diploid germ cells (i.e., primary spermatocytes and primary oocytes) go through meiosis in order to create haploid gametes (i.e., sperm and ova) for the purpose of sexual reproduction.
There are three stages of cellular interphase, with each phase ending when a cellular checkpoint checks the accuracy of the stage's completion before proceeding to the next. The stages of interphase are:
The duration of time spent in interphase and in each stage of interphase is variable and depends on both the type of cell and the species of organism it belongs to. Most cells of adult mammals spend about 24 hours in interphase; this accounts for about 90%-96% of the total time involved in cell division. Interphase includes G1, S, and G2 phases. Mitosis and cytokinesis, however, are separate from interphase.
DNA double-strand breaks can be repaired during interphase by two principal processes. The first process, non-homologous end joining (NHEJ), can join the two broken ends of DNA in the G1, S and G2 phases of interphase. The second process, homologous recombinational repair (HRR), is more accurate than NHEJ in repairing double-strand breaks. However HRR is only active during the S and G2 phases of interphase when DNA replication is either partially or fully accomplished, since HRR requires two adjacent homologous chromosomes.
When G2 is completed, the cell enters a relatively brief period of nuclear and cellular division, composed of mitosis and cytokinesis, respectively. After the successful completion of mitosis and cytokinesis, both resulting daughter cells re-enter G1 of interphase.
In the cell cycle, interphase is preceded by telophase and cytokinesis of the M phase. In alternative fashion, interphase is sometimes interrupted by G0 phase, which, in some circumstances, may then end and be followed by the remaining stages of interphase. After the successful completion of the G2 checkpoint, the final checkpoint in interphase, the cell proceeds to prophase, or in plants to preprophase, which is the first stage of mitosis.
Recent media
Recent media