
Community hub
Recent from talks
Contribute something
Nothing was collected or created yet.
Kenyapithecus
View on Wikipedia
| Kenyapithecus Temporal range:
14 Million years ago | |
|---|---|

| |
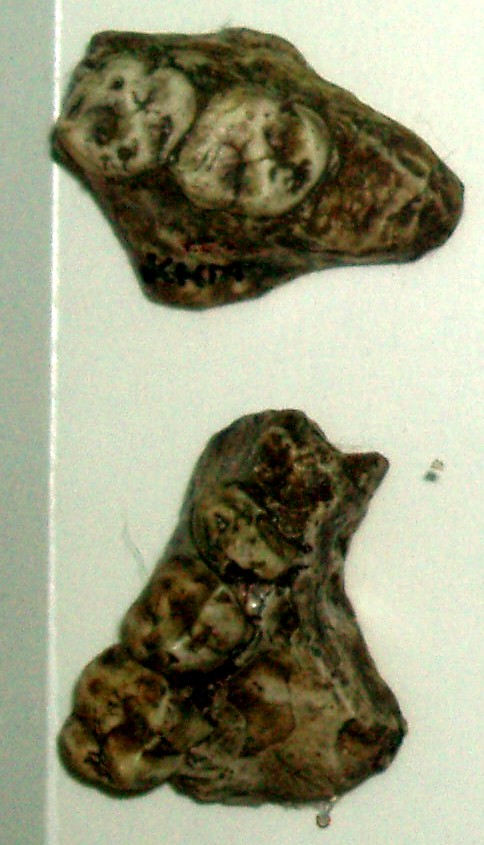
| Kenyapithecus wickeri teeth | |
| Scientific classification | |
| Kingdom: | Animalia |
| Phylum: | Chordata |
| Class: | Mammalia |
| Order: | Primates |
| Family: | Hominidae |
| Genus: | †Kenyapithecus Leakey, 1961[1] |
| Species: | †K. wickeri
|
| Binomial name | |
| †Kenyapithecus wickeri Leakey, 1961
| |
Kenyapithecus wickeri is a fossil ape discovered by Louis Leakey in 1961 at a site called Fort Ternan in Kenya. The upper jaw and teeth were dated to 14 million years ago.[2] One theory states that Kenyapithecus may be the common ancestor of all the great apes. More recent investigations suggest Kenyapithecus is more primitive than that and is only slightly more modern than Proconsul, which is considered to be an ape.
Evidence suggests that Kenyapithecus wickeri was one of the species that started a radiation of apes out of Africa.
Morphology
[edit]Impressed by Kenyapithecus's modern-looking teeth, Leakey declared Kenyapithecus to be "a very early ancestor of man himself."[3]
Kenyapithecus possessed craniodental adaptations for hard object feeding including thicker molar enamel, and a large mandible, large premolars and upper incisors that are similar to those seen in living pitheciine monkeys.[4] Kenyapithecus also possessed macaque-like limbs adapted for a knuckle-walking mode of semi-terrestrial locomotion.[5] This could show that as hominins evolved, they passed through a knuckle-walking phase.
Kenyapithecus wickeri has very distinct features, especially details in the canine teeth and is similar to modern apes.
See also
[edit]References
[edit]- ^ Ward, S.C. and Duren, D. L. (2002) "Middle and Late Miocene African Hominoids". In Hartwig, W.C. ed. The Primate Fossil Record. Cambridge University Press
- ^ L. S. B. Leakey: A new Lower Pliocene fossil primate from Kenya. In: The Annals & Magazine of Natural History, Vol. 4, Series 13, 1961, pp. 689–696
- ^ Carl Zimmer: "Kenyan Skeleton Shakes Ape Family Tree". In: Science, August 27, 1999: Vol. 285. no. 5432, pp. 1335-1337
- ^ Fleagle, J. G. (2013) Primate Adaptation and Evolution. Elsevier Academic Press
- ^ McCrossin ML, Benefit, BR Gitau, SN Palmer, AK Blue, KT. (1998) Fossil evidence for the origins of terrestriality among Old World higher primates. Primate locomotion: recent advances. New York: Plenum Press. p 353-396



