
Community hub
Recent from talks
Contribute something
Nothing was collected or created yet.
Decapodiformes
View on Wikipedia
| Decapodiformes Temporal range:
| |
|---|---|

| |
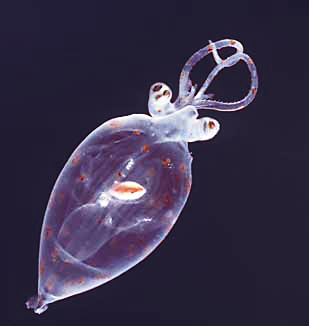
| Planktonic cranch squid paralarva; Antarctica | |
| Scientific classification | |
| Kingdom: | Animalia |
| Phylum: | Mollusca |
| Class: | Cephalopoda |
| Division: | Neocoleoidea |
| Superorder: | Decapodiformes Leach, 1817[2] |
| Synonyms | |
| |
Decapodiformes is a superorder of Cephalopoda comprising all cephalopod species with ten limbs, specifically eight short arms and two long tentacles. It is hypothesized that the ancestral coleoid had five identical pairs of limbs, and that one branch of descendants evolved a modified arm pair IV to become the Decapodiformes, while another branch of descendants evolved and then eventually lost its arm pair II, becoming the Octopodiformes.
Taxonomy
[edit]The following orders are recognised in the superorder Decapodiformes:[3]
- Bathyteuthida
- †Belemnitida
- †Diplobelida
- Idiosepida – pygmy squid
- Myopsida – coastal squid
- Oegopsida – neritic squid
- Sepiida – cuttlefish
- Sepiolida – bobtail and bottletail squid
- Spirulida – ram's horn squid
Phylogeny
[edit]Molecular analyses consistently showed that the order Teuthida is invalid as its two constituent subgroups do not form a clade that did not exclude other decapodiform orders. Thus, Teuthida is presently considered an invalid order due to lack of monophyly.[4]
Maximum Likelihood (ML) cladogram from Uribe & Zardoya 2017, based on complete mt genomes (protein-coding genes analysed at nucleotide level plus rRNA genes):[5]
The following cladogram is based on a RAxML maximum likelihood phylogram from Anderson & Lindgren 2021:[6]
| Octopodiformes |
| |||
| Decapodiformes | ||||
References
[edit]- ^ see Boletzkyida, Belemnite
- ^ Young, R. E., Vecchione, M., Mangold, K. M. (2008). Decapodiformes Leach, 1817. Squids, cuttlefishes and their relatives. in The Tree of Life Web Project
- ^ "WoRMS – World Register of Marine Species – Decapodiformes". www.marinespecies.org. Retrieved 2023-03-23.
- ^ Bieler R, Bouchet P, Gofas S, Marshall B, Rosenberg G, La Perna R, Neubauer TA, Sartori AF, Schneider S, Vos C, ter Poorten JJ, Taylor J, Dijkstra H, Finn J, Bank R, Neubert E, Moretzsohn F, Faber M, Houart R, Picton B, Garcia-Alvarez O (eds.). "Teuthida Naef, 1916". MolluscaBase. World Register of Marine Species. Retrieved 2025-07-24.
- ^ Uribe, Juan E.; Zardoya, Rafael (3 February 2017). "Revisiting the phylogeny of Cephalopoda using complete mitochondrial genomes". Journal of Molluscan Studies. 83 (2): 133–144. doi:10.1093/mollus/eyw052. hdl:10261/156228.
- ^ Anderson, Frank E.; Lindgren, Annie R. (March 2021). "Molecular Phylogenetics and Evolution Phylogenomic analyses recover a clade of large-bodied decapodiform cephalopods". Molecular Phylogenetics and Evolution. 156. doi:10.1016/j.ympev.2020.107038.
Further reading
[edit]- Young, Richard E.; Vecchione, Michael; Donovan, Desmond T. (1998). "The evolution of coleoid cephalopods and their present biodiversity and ecology". South African Journal of Marine Science. 20 (1): 393–420. doi:10.2989/025776198784126287. abstract
External links
[edit]
This cephalopod-related article is a stub. You can help Wikipedia by expanding it. |