Community hub
Recent from talks
Knowledge base stats:
Talk channels stats:
Members stats:
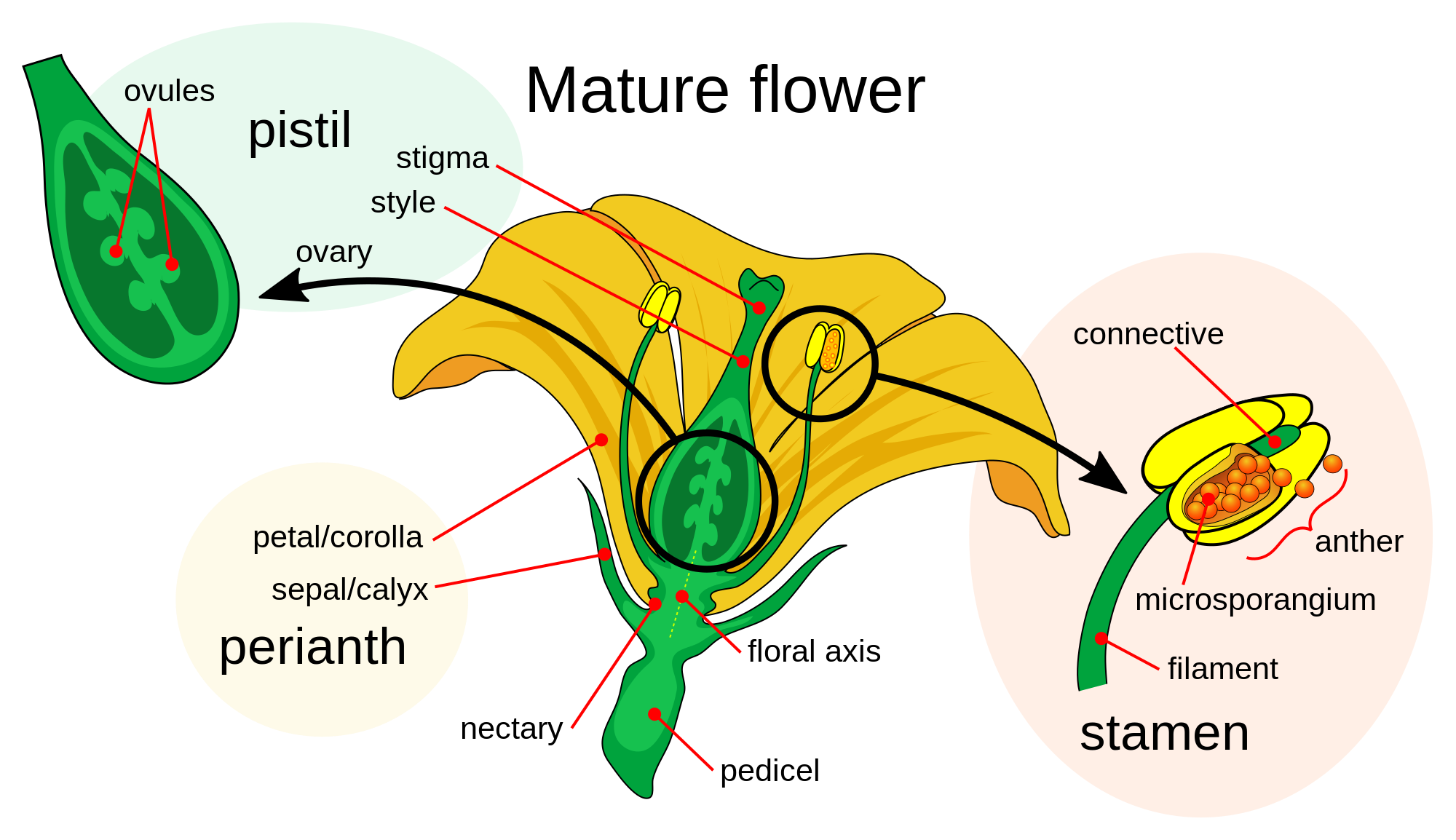
Perianth
The perianth (perigonium, perigon or perigone in monocots)[citation needed] is the non-reproductive part of a flower. It is a structure consisting of the calyx (sepals) and the corolla (petals); in perigones it consists of the tepals. It forms an envelope surrounding the sexual organs,. The term perianth is derived from Greek περί (peri, "around") and άνθος (anthos, "flower"), while perigonium is derived from περί (peri) and γόνος (gonos, "seed, sex organs"). In the mosses and liverworts (Marchantiophyta), the perianth is the sterile (neither male nor female) tube-like tissue that surrounds the female reproductive structure or developing sporophyte.
In flowering plants, the perianth may be described as being either dichlamydeous/heterochlamydeous in which the calyx and corolla are clearly separate, or homochlamydeous, in which they are indistinguishable (and the sepals and petals are collectively referred to as tepals). When the perianth is in two whorls, it is described as biseriate. While the calyx may be green, known as sepaloid, it may also be brightly coloured, and is then described as petaloid. When the undifferentiated tepals resemble petals, they are also referred to as "petaloid", as in petaloid monocots or liliod monocots, orders of monocots with brightly coloured tepals. The corolla and petals have a role in attracting pollinators, but this may be augmented by more specialised structures like the corona (see below).
When the perianth consists of separate tepals the term apotepalous is used, or syntepalous if the tepals are fused to one another. The petals may be united to form a tubular corolla (gamopetalous or sympetalous). If either the petals or sepals are entirely absent, the perianth can be described as being monochlamydeous.
Both sepals and petals may have stomata and veins, even if vestigial. In some taxa, for instance some magnolias and water lilies, the perianth is arranged in a spiral on nodes, rather than whorls. Flowers with spiral perianths tend to also be those with undifferentiated perianths.
An additional structure in some plants (e.g. Narcissus, Passiflora (passion flower), some Hippeastrum, Liliaceae) is the corona (paraperigonium, paraperigon, or paracorolla), a ring or set of appendages of adaxial tissue arising from the corolla or the outer edge of the stamens. It is often positioned where the corolla lobes arise from the corolla tube. There can be more than one corona in a flower. The milkweeds (Asclepias spp.) have three very different coronas, which collectively form a flytrap pollination scheme. Some passionflowers (Passiflora spp.) have as many as eight coronas arranged in concentric whorls.
The pappus of Asteraceae, considered to be a modified calyx, is also called a corona if it is shaped like a crown.
Hub AI
Perianth AI simulator
(@Perianth_simulator)
Perianth
The perianth (perigonium, perigon or perigone in monocots)[citation needed] is the non-reproductive part of a flower. It is a structure consisting of the calyx (sepals) and the corolla (petals); in perigones it consists of the tepals. It forms an envelope surrounding the sexual organs,. The term perianth is derived from Greek περί (peri, "around") and άνθος (anthos, "flower"), while perigonium is derived from περί (peri) and γόνος (gonos, "seed, sex organs"). In the mosses and liverworts (Marchantiophyta), the perianth is the sterile (neither male nor female) tube-like tissue that surrounds the female reproductive structure or developing sporophyte.
In flowering plants, the perianth may be described as being either dichlamydeous/heterochlamydeous in which the calyx and corolla are clearly separate, or homochlamydeous, in which they are indistinguishable (and the sepals and petals are collectively referred to as tepals). When the perianth is in two whorls, it is described as biseriate. While the calyx may be green, known as sepaloid, it may also be brightly coloured, and is then described as petaloid. When the undifferentiated tepals resemble petals, they are also referred to as "petaloid", as in petaloid monocots or liliod monocots, orders of monocots with brightly coloured tepals. The corolla and petals have a role in attracting pollinators, but this may be augmented by more specialised structures like the corona (see below).
When the perianth consists of separate tepals the term apotepalous is used, or syntepalous if the tepals are fused to one another. The petals may be united to form a tubular corolla (gamopetalous or sympetalous). If either the petals or sepals are entirely absent, the perianth can be described as being monochlamydeous.
Both sepals and petals may have stomata and veins, even if vestigial. In some taxa, for instance some magnolias and water lilies, the perianth is arranged in a spiral on nodes, rather than whorls. Flowers with spiral perianths tend to also be those with undifferentiated perianths.
An additional structure in some plants (e.g. Narcissus, Passiflora (passion flower), some Hippeastrum, Liliaceae) is the corona (paraperigonium, paraperigon, or paracorolla), a ring or set of appendages of adaxial tissue arising from the corolla or the outer edge of the stamens. It is often positioned where the corolla lobes arise from the corolla tube. There can be more than one corona in a flower. The milkweeds (Asclepias spp.) have three very different coronas, which collectively form a flytrap pollination scheme. Some passionflowers (Passiflora spp.) have as many as eight coronas arranged in concentric whorls.
The pappus of Asteraceae, considered to be a modified calyx, is also called a corona if it is shaped like a crown.