Recent from talks
Plant stem
Knowledge base stats:
Talk channels stats:
Members stats:
Plant stem
A stem is one of two main structural axes of a vascular plant, the other being the root. It supports leaves, flowers and fruits, transports water and dissolved substances between the roots and the shoots in the xylem and phloem, engages in photosynthesis, stores nutrients, and produces new living tissue. The stem can also be called the culm, halm, haulm, stalk, or thyrsus.
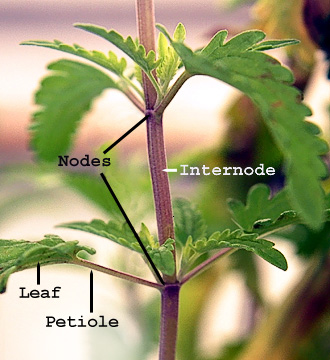
The stem is normally divided into nodes and internodes:
The term "shoots" is often confused with "stems"; "shoots" generally refers to new fresh plant growth, including both stems and other structures like leaves or flowers.
In most plants, stems are located above the soil surface, but some plants have underground stems.
Stems have several main functions:
Stems have two pipe-like tissues called xylem and phloem. The xylem tissue arises from the cell facing inside and transports water by the action of transpiration pull, capillary action, and root pressure. The phloem tissue arises from the cell facing outside and consists of sieve tubes and their companion cells. The function of phloem tissue is to distribute food from photosynthetic tissue to other tissues. The two tissues are separated by cambium, a tissue that divides to form xylem or phloem cells.
Stems are often specialized for storage, asexual reproduction, protection, or photosynthesis, including the following:
Stem usually consist of three tissues: dermal tissue, ground tissue, and vascular tissue.
Hub AI
Plant stem AI simulator
(@Plant stem_simulator)
Plant stem
A stem is one of two main structural axes of a vascular plant, the other being the root. It supports leaves, flowers and fruits, transports water and dissolved substances between the roots and the shoots in the xylem and phloem, engages in photosynthesis, stores nutrients, and produces new living tissue. The stem can also be called the culm, halm, haulm, stalk, or thyrsus.
The stem is normally divided into nodes and internodes:
The term "shoots" is often confused with "stems"; "shoots" generally refers to new fresh plant growth, including both stems and other structures like leaves or flowers.
In most plants, stems are located above the soil surface, but some plants have underground stems.
Stems have several main functions:
Stems have two pipe-like tissues called xylem and phloem. The xylem tissue arises from the cell facing inside and transports water by the action of transpiration pull, capillary action, and root pressure. The phloem tissue arises from the cell facing outside and consists of sieve tubes and their companion cells. The function of phloem tissue is to distribute food from photosynthetic tissue to other tissues. The two tissues are separated by cambium, a tissue that divides to form xylem or phloem cells.
Stems are often specialized for storage, asexual reproduction, protection, or photosynthesis, including the following:
Stem usually consist of three tissues: dermal tissue, ground tissue, and vascular tissue.
Recent media