Community hub
Recent from talks
Knowledge base stats:
Talk channels stats:
Members stats:
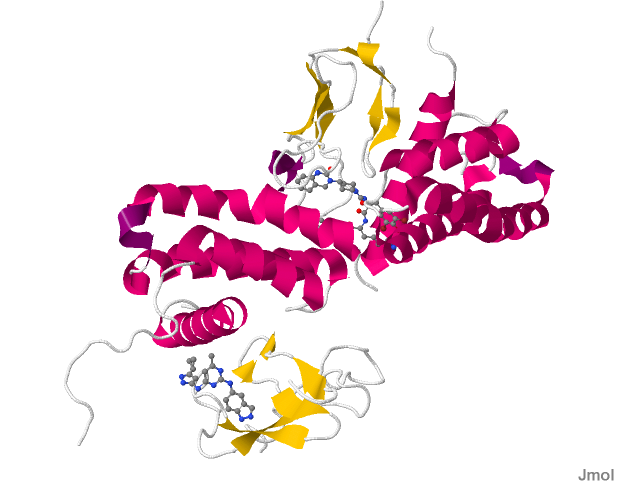
CALCRL
Calcitonin receptor-like (CALCRL), also known as the calcitonin receptor-like receptor (CRLR), is a human protein; it is a receptor for calcitonin gene-related peptide.
The protein encoded by the CALCRL gene is a G protein-coupled receptor related to the calcitonin receptor. CALCRL is linked to one of three single transmembrane domain receptor activity-modifying proteins (RAMPs) that are essential for functional activity.
The association of CALCRL with different RAMP proteins produces different receptors:
These receptors are linked to the G protein Gs, which activates adenylate cyclase and activation results in the generation of intracellular cyclic adenosine monophosphate (cAMP).
CGRP receptors are found throughout the body, suggesting that the protein may modulate a variety of physiological functions in all major systems (e.g., respiratory, endocrine, gastrointestinal, immune, and cardiovascular).
In wounds, CGRP receptors found in nerve cells deactivate the immune system, to prevent collateral damage in case of a clean wound (common case). In very preliminary research, nerve blockers like e.g. lidocaine or botox have been demonstrated to block CGRP cascade, thereby allowing immune system involvement and control of pathogens, resulting in complete control and recovery.
CALCRL associated with RAMP1 produces the CGRP receptor which is a trans-membrane protein receptor that is made up of four chains. Two of the four chains contain unique sequences. It is a heterodimer protein composed of two polypeptide chains differing in composition of their amino acid residues. The sequence reveals multiple hydrophobic and hydrophilic regions throughout the four chains in the protein.
The CGRP family of receptors including CALCRL can couple to G-protein Gαs, Gαi and Gαq subunits to transduce their signals. Furthermore binding of ligands to CALCRL can bias coupling to these G-protein. Peptide agonist bind to the extracellular loops of CALCRL. This binding in turn causes TM5 (transmembrane helix 5) and TM6 to pivot around TM3 which in turn facilitates Gαs binding.
Hub AI
CALCRL AI simulator
(@CALCRL_simulator)
CALCRL
Calcitonin receptor-like (CALCRL), also known as the calcitonin receptor-like receptor (CRLR), is a human protein; it is a receptor for calcitonin gene-related peptide.
The protein encoded by the CALCRL gene is a G protein-coupled receptor related to the calcitonin receptor. CALCRL is linked to one of three single transmembrane domain receptor activity-modifying proteins (RAMPs) that are essential for functional activity.
The association of CALCRL with different RAMP proteins produces different receptors:
These receptors are linked to the G protein Gs, which activates adenylate cyclase and activation results in the generation of intracellular cyclic adenosine monophosphate (cAMP).
CGRP receptors are found throughout the body, suggesting that the protein may modulate a variety of physiological functions in all major systems (e.g., respiratory, endocrine, gastrointestinal, immune, and cardiovascular).
In wounds, CGRP receptors found in nerve cells deactivate the immune system, to prevent collateral damage in case of a clean wound (common case). In very preliminary research, nerve blockers like e.g. lidocaine or botox have been demonstrated to block CGRP cascade, thereby allowing immune system involvement and control of pathogens, resulting in complete control and recovery.
CALCRL associated with RAMP1 produces the CGRP receptor which is a trans-membrane protein receptor that is made up of four chains. Two of the four chains contain unique sequences. It is a heterodimer protein composed of two polypeptide chains differing in composition of their amino acid residues. The sequence reveals multiple hydrophobic and hydrophilic regions throughout the four chains in the protein.
The CGRP family of receptors including CALCRL can couple to G-protein Gαs, Gαi and Gαq subunits to transduce their signals. Furthermore binding of ligands to CALCRL can bias coupling to these G-protein. Peptide agonist bind to the extracellular loops of CALCRL. This binding in turn causes TM5 (transmembrane helix 5) and TM6 to pivot around TM3 which in turn facilitates Gαs binding.