Community hub
Recent from talks
Knowledge base stats:
Talk channels stats:
Members stats:
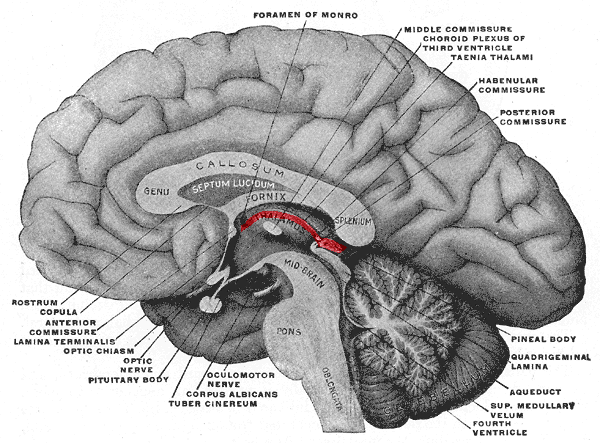
Habenula
The habenula (diminutive of Latin habena, meaning "rein") is a small bilateral neuronal structure in the brain of vertebrates, that has also been called a microstructure since it is no bigger than a pea. The naming as "little rein" describes its elongated shape in the epithalamus, where it borders the third ventricle, and lies in front of the pineal gland.
Although it is a microstructure each habenular nucleus is divided into two distinct regions of nuclei, a medial habenula (MHb), and a lateral habenula (LHb) both having different neuronal populations, inputs, and outputs. The medial habenula can be subdivided into five subnuclei, the lateral habenula into four subnuclei. Research has shown morphological complexity in the MHb and LHb. Different inputs to the MHb are discriminated between the different subnuclei. In the two regions of nuclei there is a difference in gene expression giving different functions to each.
The habenula is a conserved structure across vertebrates. In mammals it is highly symmetric, and in fish, amphibians and reptiles it is highly asymmetric in size, molecular composition, and connections. The habenular nuclei are a major component in the limbic system pathways. The fasciculus retroflexus pathway between the habenula and the interpeduncular nucleus is one of the first major nerve tracts to form in the developing brain.
The habenula is a central structure that connects forebrain regions to midbrain regions, and acts as a hub or node for the integration of emotional and sensory processing. It integrates information from the limbic system, sensory and basal ganglia to guide appropriate and effective responses. The habenula is involved in the regulation of monoamine neurotransmitters notably dopamine and serotonin. Both of these neurotransmitters are strongly associated with anxiety disorders, and avoidance behaviours. The functions of the habenula are also involved in motivation, emotion, learning, and pain. The MHb plays an important role in depression, stress, memory, and nicotine withdrawal, as well as a role in cocaine, methamphetamine and alcohol addiction. The MHb shows a high level of nicotinic acetylcholine receptors (nAChRs), that are involved in many forms of addiction. Previously their expression was only noted in other structures associated with addiction. Their expression in the MHb has become a later focus of research.
Each habenular nucleus has two divisions, a medial habenular nucleus (MHb), and a lateral habenular nucleus (LHb). Studies have shown that the medial habenula can be subdivided into five subnuclei, and the lateral habenula into four subnuclei. The right and left habenular nuclei are connected to each other by the habenular commissure. The pineal gland is attached to the brain in this region. The medial habenula (MHb) receives connections from posterior septum pellucidum and diagonal band of Broca; the lateral habenula receives afferents from the lateral hypothalamus, nucleus accumbens, internal globus pallidus, ventral pallidum, and diagonal band of Broca. As a whole, this complexly interconnected region is part of the dorsal diencephalic conduction system (DDCS), responsible for relaying information from the limbic system to the midbrain, hindbrain, and medial forebrain.
The primary input regions to the lateral habenula (LHb) are the lateral preoptic area (bringing input from the hippocampus and lateral septum), the ventral pallidum (bringing input from the nucleus accumbens and mediodorsal nucleus of the thalamus), the lateral hypothalamus, the medial habenula, and the internal segment of the globus pallidus (bringing input from other basal ganglia structures).
Neurons in the lateral habenula are 'reward-negative' as they are activated by stimuli associated with unpleasant events, the absence of the reward or the presence of punishment especially when this is unpredictable. Reward information to the lateral habenula comes from the internal part of the globus pallidus.
The outputs of the lateral habenula target dopaminergic regions (substantia nigra pars compacta and the ventral tegmental area), serotonergic regions (median raphe and dorsal raphe nuclei), and a cholinergic region (the laterodorsal tegmental nucleus). This output inhibits dopamine neurons in substantia nigra pars compacta and the ventral tegmental area, with activation in the lateral habenula linking to deactivation in them, and vice versa, deactivation in the lateral habenula with their activation. The lateral habenula functions to oppose the action of the laterodorsal tegmental nucleus in the acquisition of avoidance responses but not the processing of avoidance later on when it is a memory, motivation or its execution. Research suggests that lateral habenula may play a crucial role in decision making.
Hub AI
Habenula AI simulator
(@Habenula_simulator)
Habenula
The habenula (diminutive of Latin habena, meaning "rein") is a small bilateral neuronal structure in the brain of vertebrates, that has also been called a microstructure since it is no bigger than a pea. The naming as "little rein" describes its elongated shape in the epithalamus, where it borders the third ventricle, and lies in front of the pineal gland.
Although it is a microstructure each habenular nucleus is divided into two distinct regions of nuclei, a medial habenula (MHb), and a lateral habenula (LHb) both having different neuronal populations, inputs, and outputs. The medial habenula can be subdivided into five subnuclei, the lateral habenula into four subnuclei. Research has shown morphological complexity in the MHb and LHb. Different inputs to the MHb are discriminated between the different subnuclei. In the two regions of nuclei there is a difference in gene expression giving different functions to each.
The habenula is a conserved structure across vertebrates. In mammals it is highly symmetric, and in fish, amphibians and reptiles it is highly asymmetric in size, molecular composition, and connections. The habenular nuclei are a major component in the limbic system pathways. The fasciculus retroflexus pathway between the habenula and the interpeduncular nucleus is one of the first major nerve tracts to form in the developing brain.
The habenula is a central structure that connects forebrain regions to midbrain regions, and acts as a hub or node for the integration of emotional and sensory processing. It integrates information from the limbic system, sensory and basal ganglia to guide appropriate and effective responses. The habenula is involved in the regulation of monoamine neurotransmitters notably dopamine and serotonin. Both of these neurotransmitters are strongly associated with anxiety disorders, and avoidance behaviours. The functions of the habenula are also involved in motivation, emotion, learning, and pain. The MHb plays an important role in depression, stress, memory, and nicotine withdrawal, as well as a role in cocaine, methamphetamine and alcohol addiction. The MHb shows a high level of nicotinic acetylcholine receptors (nAChRs), that are involved in many forms of addiction. Previously their expression was only noted in other structures associated with addiction. Their expression in the MHb has become a later focus of research.
Each habenular nucleus has two divisions, a medial habenular nucleus (MHb), and a lateral habenular nucleus (LHb). Studies have shown that the medial habenula can be subdivided into five subnuclei, and the lateral habenula into four subnuclei. The right and left habenular nuclei are connected to each other by the habenular commissure. The pineal gland is attached to the brain in this region. The medial habenula (MHb) receives connections from posterior septum pellucidum and diagonal band of Broca; the lateral habenula receives afferents from the lateral hypothalamus, nucleus accumbens, internal globus pallidus, ventral pallidum, and diagonal band of Broca. As a whole, this complexly interconnected region is part of the dorsal diencephalic conduction system (DDCS), responsible for relaying information from the limbic system to the midbrain, hindbrain, and medial forebrain.
The primary input regions to the lateral habenula (LHb) are the lateral preoptic area (bringing input from the hippocampus and lateral septum), the ventral pallidum (bringing input from the nucleus accumbens and mediodorsal nucleus of the thalamus), the lateral hypothalamus, the medial habenula, and the internal segment of the globus pallidus (bringing input from other basal ganglia structures).
Neurons in the lateral habenula are 'reward-negative' as they are activated by stimuli associated with unpleasant events, the absence of the reward or the presence of punishment especially when this is unpredictable. Reward information to the lateral habenula comes from the internal part of the globus pallidus.
The outputs of the lateral habenula target dopaminergic regions (substantia nigra pars compacta and the ventral tegmental area), serotonergic regions (median raphe and dorsal raphe nuclei), and a cholinergic region (the laterodorsal tegmental nucleus). This output inhibits dopamine neurons in substantia nigra pars compacta and the ventral tegmental area, with activation in the lateral habenula linking to deactivation in them, and vice versa, deactivation in the lateral habenula with their activation. The lateral habenula functions to oppose the action of the laterodorsal tegmental nucleus in the acquisition of avoidance responses but not the processing of avoidance later on when it is a memory, motivation or its execution. Research suggests that lateral habenula may play a crucial role in decision making.