Recent from talks
Taste
Knowledge base stats:
Talk channels stats:
Members stats:
Taste
The gustatory system or sense of taste is the sensory system that is partially responsible for the perception of taste. Taste is the perception stimulated when a substance in the mouth reacts chemically with taste receptor cells located on taste buds in the oral cavity, mostly on the tongue. Taste, along with the sense of smell and trigeminal nerve stimulation (registering texture, pain, and temperature), determines flavors of food and other substances. Humans have taste receptors on taste buds and other areas, including the upper surface of the tongue and the epiglottis. The gustatory cortex is responsible for the perception of taste.
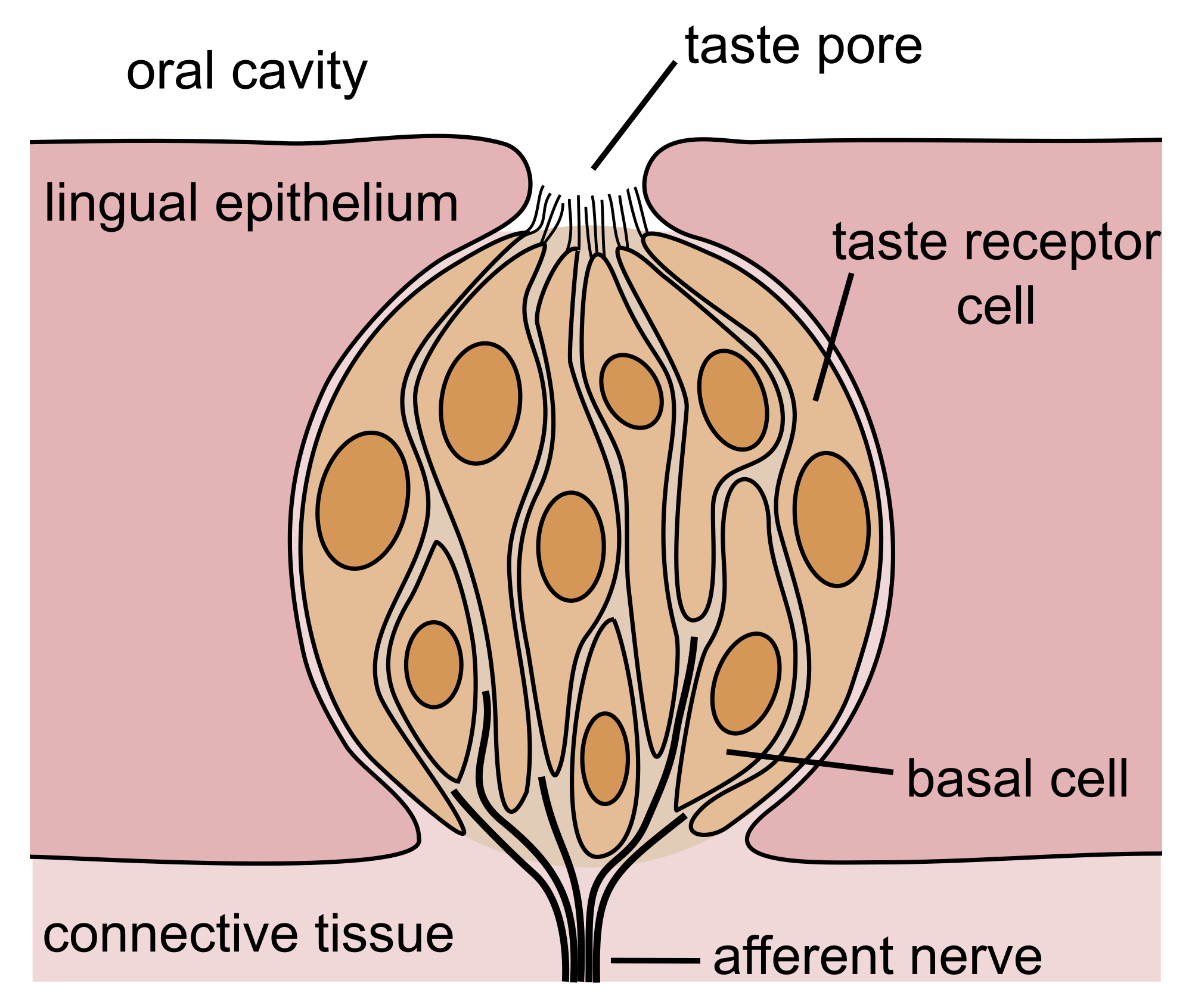
The tongue is covered with thousands of small bumps called papillae, which are visible to the naked eye. Within each papilla are hundreds of taste buds. The exceptions to this is the filiform papillae that do not contain taste buds. There are between 2000 and 5000 taste buds that are located on the back and front of the tongue. Others are located on the roof, sides and back of the mouth, and in the throat. Each taste bud contains 50 to 100 taste receptor cells.
Taste receptors in the mouth sense the five basic tastes: sweetness, sourness, saltiness, bitterness, and savoriness (also known as savory or umami). Scientific experiments have demonstrated that these five tastes exist and are distinct from one another. Taste buds are able to tell different tastes apart when they interact with different molecules or ions. Sweetness, savoriness, and bitter tastes are triggered by the binding of molecules to G protein-coupled receptors on the cell membranes of taste buds. Saltiness and sourness are perceived when alkali metals or hydrogen ions meet taste buds, respectively.
The basic tastes contribute only partially to the sensation and flavor of food in the mouth—other factors include smell, detected by the olfactory epithelium of the nose; texture, detected through a variety of mechanoreceptors, muscle nerves, etc.; temperature, detected by temperature receptors; and "coolness" (such as of menthol) and "hotness" (pungency), by chemesthesis.
As the gustatory system senses both harmful and beneficial things, all basic tastes bring either caution or craving depending upon the effect the things they sense have on the body. Sweetness helps to identify energy-rich foods, while bitterness warns people of poisons.
Among humans, taste perception begins to fade during ageing, tongue papillae are lost, and saliva production slowly decreases. Humans can also have distortion of tastes (dysgeusia). Not all mammals share the same tastes: some rodents can taste starch (which humans cannot), cats cannot taste sweetness, and several other carnivores, including hyenas, dolphins, and sea lions, have lost the ability to sense up to four of their ancestral five basic tastes.
The gustatory system allows animals to distinguish between safe and harmful food and to gauge different foods' nutritional value. Digestive enzymes in saliva begin to dissolve food into base chemicals that are washed over the papillae and detected as tastes by the taste buds. The tongue is covered with thousands of small bumps called papillae, which are visible to the naked eye. Within each papilla are hundreds of taste buds. The exception to this are the filiform papillae, which do not contain taste buds. There are between 2,000 and 5,000 taste buds that are located on the back and front of the tongue. Others are located on the roof, sides and back of the mouth, and in the throat. Each taste bud contains 50 to 100 taste-receptor cells.
The five specific tastes received by taste receptors are saltiness, sweetness, bitterness, sourness, and savoriness (often known by its Japanese name umami, which translates to 'deliciousness').
Hub AI
Taste AI simulator
(@Taste_simulator)
Taste
The gustatory system or sense of taste is the sensory system that is partially responsible for the perception of taste. Taste is the perception stimulated when a substance in the mouth reacts chemically with taste receptor cells located on taste buds in the oral cavity, mostly on the tongue. Taste, along with the sense of smell and trigeminal nerve stimulation (registering texture, pain, and temperature), determines flavors of food and other substances. Humans have taste receptors on taste buds and other areas, including the upper surface of the tongue and the epiglottis. The gustatory cortex is responsible for the perception of taste.
The tongue is covered with thousands of small bumps called papillae, which are visible to the naked eye. Within each papilla are hundreds of taste buds. The exceptions to this is the filiform papillae that do not contain taste buds. There are between 2000 and 5000 taste buds that are located on the back and front of the tongue. Others are located on the roof, sides and back of the mouth, and in the throat. Each taste bud contains 50 to 100 taste receptor cells.
Taste receptors in the mouth sense the five basic tastes: sweetness, sourness, saltiness, bitterness, and savoriness (also known as savory or umami). Scientific experiments have demonstrated that these five tastes exist and are distinct from one another. Taste buds are able to tell different tastes apart when they interact with different molecules or ions. Sweetness, savoriness, and bitter tastes are triggered by the binding of molecules to G protein-coupled receptors on the cell membranes of taste buds. Saltiness and sourness are perceived when alkali metals or hydrogen ions meet taste buds, respectively.
The basic tastes contribute only partially to the sensation and flavor of food in the mouth—other factors include smell, detected by the olfactory epithelium of the nose; texture, detected through a variety of mechanoreceptors, muscle nerves, etc.; temperature, detected by temperature receptors; and "coolness" (such as of menthol) and "hotness" (pungency), by chemesthesis.
As the gustatory system senses both harmful and beneficial things, all basic tastes bring either caution or craving depending upon the effect the things they sense have on the body. Sweetness helps to identify energy-rich foods, while bitterness warns people of poisons.
Among humans, taste perception begins to fade during ageing, tongue papillae are lost, and saliva production slowly decreases. Humans can also have distortion of tastes (dysgeusia). Not all mammals share the same tastes: some rodents can taste starch (which humans cannot), cats cannot taste sweetness, and several other carnivores, including hyenas, dolphins, and sea lions, have lost the ability to sense up to four of their ancestral five basic tastes.
The gustatory system allows animals to distinguish between safe and harmful food and to gauge different foods' nutritional value. Digestive enzymes in saliva begin to dissolve food into base chemicals that are washed over the papillae and detected as tastes by the taste buds. The tongue is covered with thousands of small bumps called papillae, which are visible to the naked eye. Within each papilla are hundreds of taste buds. The exception to this are the filiform papillae, which do not contain taste buds. There are between 2,000 and 5,000 taste buds that are located on the back and front of the tongue. Others are located on the roof, sides and back of the mouth, and in the throat. Each taste bud contains 50 to 100 taste-receptor cells.
The five specific tastes received by taste receptors are saltiness, sweetness, bitterness, sourness, and savoriness (often known by its Japanese name umami, which translates to 'deliciousness').
Recent media