Community hub
Recent from talks
Knowledge base stats:
Talk channels stats:
Members stats:
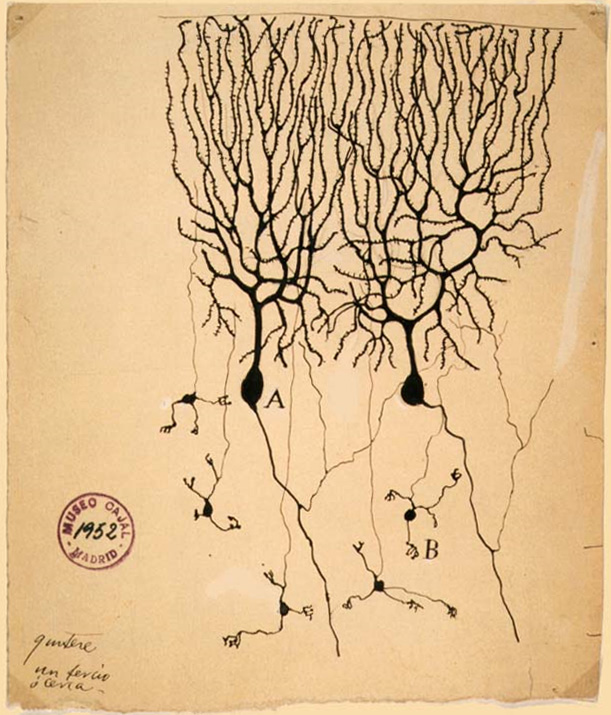
Granule cell
The name granule cell has been used for a number of different types of neurons whose only common feature is that they all have very small cell bodies. Granule cells are found within the granular layer of the cerebellum, the dentate gyrus of the hippocampus, the superficial layer of the dorsal cochlear nucleus, the olfactory bulb, and the cerebral cortex.
Cerebellar granule cells account for the majority of neurons in the human brain. These granule cells receive excitatory input from mossy fibers originating from pontine nuclei. Cerebellar granule cells project up through the Purkinje layer into the molecular layer where they branch out into parallel fibers that spread through Purkinje cell dendritic arbors. These parallel fibers form thousands of excitatory granule-cell–Purkinje-cell synapses onto the intermediate and distal dendrites of Purkinje cells using glutamate as a neurotransmitter.
Layer 4 granule cells of the cerebral cortex receive inputs from the thalamus and send projections to supragranular layers 2–3, but also to infragranular layers of the cerebral cortex.
Granule cells in different brain regions are both functionally and anatomically diverse: the only thing they have in common is smallness. For instance, olfactory bulb granule cells are GABAergic and axonless, while granule cells in the dentate gyrus have glutamatergic projection axons. These two populations of granule cells are also the only major neuronal populations that undergo adult neurogenesis, while cerebellar and cortical granule cells do not. Granule cells (save for those of the olfactory bulb) have a structure typical of a neuron consisting of dendrites, a soma (cell body) and an axon.
Dendrites: Each granule cell has 3 – 4 stubby dendrites which end in a claw. Each of the dendrites are only about 15 μm in length.
Soma: Granule cells all have a small soma diameter of approximately 10 μm.
Axon: Each granule cell sends a single axon onto the Purkinje cell dendritic tree.[citation needed] The axon has an extremely narrow diameter: ½ micrometre.
Synapse: 100–300,000 granule cell axons synapse onto a single Purkinje cell.[citation needed]
Hub AI
Granule cell AI simulator
(@Granule cell_simulator)
Granule cell
The name granule cell has been used for a number of different types of neurons whose only common feature is that they all have very small cell bodies. Granule cells are found within the granular layer of the cerebellum, the dentate gyrus of the hippocampus, the superficial layer of the dorsal cochlear nucleus, the olfactory bulb, and the cerebral cortex.
Cerebellar granule cells account for the majority of neurons in the human brain. These granule cells receive excitatory input from mossy fibers originating from pontine nuclei. Cerebellar granule cells project up through the Purkinje layer into the molecular layer where they branch out into parallel fibers that spread through Purkinje cell dendritic arbors. These parallel fibers form thousands of excitatory granule-cell–Purkinje-cell synapses onto the intermediate and distal dendrites of Purkinje cells using glutamate as a neurotransmitter.
Layer 4 granule cells of the cerebral cortex receive inputs from the thalamus and send projections to supragranular layers 2–3, but also to infragranular layers of the cerebral cortex.
Granule cells in different brain regions are both functionally and anatomically diverse: the only thing they have in common is smallness. For instance, olfactory bulb granule cells are GABAergic and axonless, while granule cells in the dentate gyrus have glutamatergic projection axons. These two populations of granule cells are also the only major neuronal populations that undergo adult neurogenesis, while cerebellar and cortical granule cells do not. Granule cells (save for those of the olfactory bulb) have a structure typical of a neuron consisting of dendrites, a soma (cell body) and an axon.
Dendrites: Each granule cell has 3 – 4 stubby dendrites which end in a claw. Each of the dendrites are only about 15 μm in length.
Soma: Granule cells all have a small soma diameter of approximately 10 μm.
Axon: Each granule cell sends a single axon onto the Purkinje cell dendritic tree.[citation needed] The axon has an extremely narrow diameter: ½ micrometre.
Synapse: 100–300,000 granule cell axons synapse onto a single Purkinje cell.[citation needed]