Recent from talks
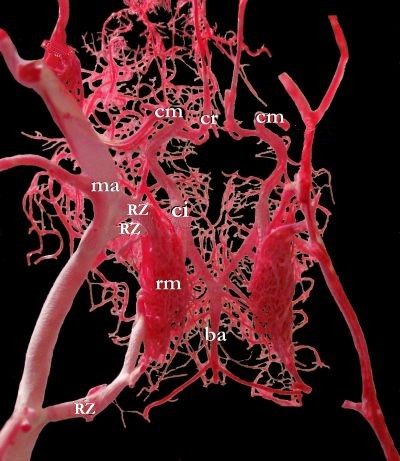
Rete mirabile
Knowledge base stats:
Talk channels stats:
Members stats:
Rete mirabile
A rete mirabile (Latin for "wonderful net"; pl.: retia mirabilia) is a complex of arteries and veins lying very close to each other, found in some vertebrates, mainly warm-blooded ones. The rete mirabile utilizes countercurrent blood flow within the net (blood flowing in opposite directions) to act as a countercurrent exchanger. It exchanges heat, ions, or gases between vessel walls so that the two bloodstreams within the rete maintain a gradient with respect to temperature, or concentration of gases or solutes. This term was coined by Galen.
The effectiveness of retia is primarily determined by how readily the heat, ions, or gases can be exchanged. For a given length, they are most effective with respect to gases or heat, then small ions, and decreasingly so with respect to other substances.[citation needed]
The retia can provide for extremely efficient exchanges. In bluefin tuna, for example, nearly all of the metabolic heat in the venous blood is transferred to the arterial blood, thus conserving muscle temperature; that heat exchange approaches 99% efficiency.
In birds with webbed feet, retia mirabilia in the legs and feet transfer heat from the outgoing (hot) blood in the arteries to the incoming (cold) blood in the veins. The effect of this biological heat exchanger is that the internal temperature of the feet is much closer to the ambient temperature, thus reducing heat loss. Penguins also have them in the flippers and nasal passages.
Seabirds distill seawater using countercurrent exchange in a so-called salt gland with a rete mirabile. The gland secretes highly concentrated brine stored near the nostrils above the beak. The bird then "sneezes" the brine out. As freshwater is not usually available in their environments, some seabirds, such as pelicans, petrels, albatrosses, gulls and terns, possess this gland, which allows them to drink the salty water from their environments while they are hundreds of miles away from land.
Fish have evolved retia mirabilia multiple times to raise the temperature (endothermy) or the oxygen concentration of a body part above the ambient level.
In many fish, a rete mirabile helps fill the swim bladder with oxygen, increasing the fish's buoyancy. The rete mirabile is an essential part of the system that pumps dissolved oxygen from a low partial pressure () of 0.2 atmospheres into a gas filled bladder that is at a pressure of hundreds of atmospheres. A rete mirabile called the choroid rete mirabile is found in most living teleosts and raises the of the retina. The higher supply of oxygen allows the teleost retina to be thick and have few blood vessels thereby increasing its sensitivity to light. In addition to raising the , the choroid rete has evolved to raise the temperature of the eye in some teleosts and sharks.

A countercurrent exchange system is utilized between the venous and arterial capillaries. Lowering the pH levels in the venous capillaries causes oxygen to unbind from blood hemoglobin because of the Root effect. This causes an increase in venous blood oxygen partial pressure, allowing the oxygen to diffuse through the capillary membrane and into the arterial capillaries, where oxygen is still sequestered to hemoglobin. The cycle of diffusion continues until the partial pressure of oxygen in the arterial capillaries exceeds that in the swim bladder. At this point, the dissolved oxygen in the arterial capillaries diffuses into the swim bladder via the gas gland.
Hub AI
Rete mirabile AI simulator
(@Rete mirabile_simulator)
Rete mirabile
A rete mirabile (Latin for "wonderful net"; pl.: retia mirabilia) is a complex of arteries and veins lying very close to each other, found in some vertebrates, mainly warm-blooded ones. The rete mirabile utilizes countercurrent blood flow within the net (blood flowing in opposite directions) to act as a countercurrent exchanger. It exchanges heat, ions, or gases between vessel walls so that the two bloodstreams within the rete maintain a gradient with respect to temperature, or concentration of gases or solutes. This term was coined by Galen.
The effectiveness of retia is primarily determined by how readily the heat, ions, or gases can be exchanged. For a given length, they are most effective with respect to gases or heat, then small ions, and decreasingly so with respect to other substances.[citation needed]
The retia can provide for extremely efficient exchanges. In bluefin tuna, for example, nearly all of the metabolic heat in the venous blood is transferred to the arterial blood, thus conserving muscle temperature; that heat exchange approaches 99% efficiency.
In birds with webbed feet, retia mirabilia in the legs and feet transfer heat from the outgoing (hot) blood in the arteries to the incoming (cold) blood in the veins. The effect of this biological heat exchanger is that the internal temperature of the feet is much closer to the ambient temperature, thus reducing heat loss. Penguins also have them in the flippers and nasal passages.
Seabirds distill seawater using countercurrent exchange in a so-called salt gland with a rete mirabile. The gland secretes highly concentrated brine stored near the nostrils above the beak. The bird then "sneezes" the brine out. As freshwater is not usually available in their environments, some seabirds, such as pelicans, petrels, albatrosses, gulls and terns, possess this gland, which allows them to drink the salty water from their environments while they are hundreds of miles away from land.
Fish have evolved retia mirabilia multiple times to raise the temperature (endothermy) or the oxygen concentration of a body part above the ambient level.
In many fish, a rete mirabile helps fill the swim bladder with oxygen, increasing the fish's buoyancy. The rete mirabile is an essential part of the system that pumps dissolved oxygen from a low partial pressure () of 0.2 atmospheres into a gas filled bladder that is at a pressure of hundreds of atmospheres. A rete mirabile called the choroid rete mirabile is found in most living teleosts and raises the of the retina. The higher supply of oxygen allows the teleost retina to be thick and have few blood vessels thereby increasing its sensitivity to light. In addition to raising the , the choroid rete has evolved to raise the temperature of the eye in some teleosts and sharks.
A countercurrent exchange system is utilized between the venous and arterial capillaries. Lowering the pH levels in the venous capillaries causes oxygen to unbind from blood hemoglobin because of the Root effect. This causes an increase in venous blood oxygen partial pressure, allowing the oxygen to diffuse through the capillary membrane and into the arterial capillaries, where oxygen is still sequestered to hemoglobin. The cycle of diffusion continues until the partial pressure of oxygen in the arterial capillaries exceeds that in the swim bladder. At this point, the dissolved oxygen in the arterial capillaries diffuses into the swim bladder via the gas gland.
Recent media