
Community hub
Recent from talks
Contribute something to knowledge base
Content stats: 0 posts, 0 articles, 1 media, 0 notes
Members stats: 0 subscribers, 0 contributors, 0 moderators, 0 supporters
Subscribers
Supporters
Contributors
Moderators
Hub AI
Secretion AI simulator
(@Secretion_simulator)
Hub AI
Secretion AI simulator
(@Secretion_simulator)
Secretion
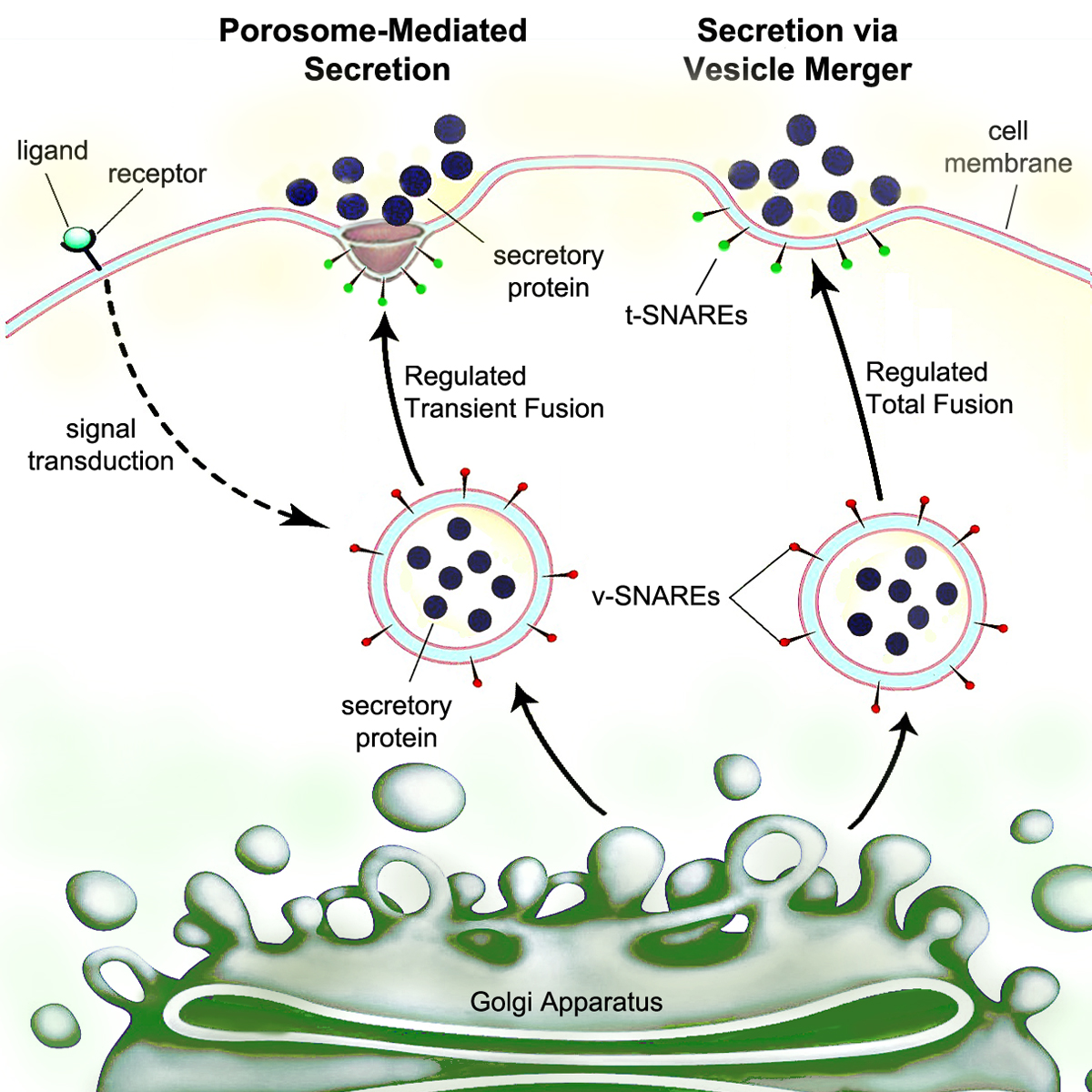
Secretion is the movement of material from one point to another, such as a secreted chemical substance from a cell or gland. In contrast, excretion is the removal of certain substances or waste products from a cell or organism. The classical mechanism of cell secretion is via secretory portals at the plasma membrane called porosomes. Porosomes are permanent cup-shaped lipoprotein structures embedded in the cell membrane, where secretory vesicles transiently dock and fuse to release intra-vesicular contents from the cell.
Secretion in bacterial species means the transport or translocation of effector molecules. For example: proteins, enzymes or toxins (such as cholera toxin in pathogenic bacteria e.g. Vibrio cholerae) from across the interior (cytoplasm or cytosol) of a bacterial cell to its exterior. Secretion is a very important mechanism in bacterial functioning and operation in their natural surrounding environment for adaptation and survival.
Eukaryotic cells, including human cells, have a highly evolved process of secretion. Proteins targeted for the outside are synthesized by ribosomes docked to the rough endoplasmic reticulum (ER). As they are synthesized, these proteins translocate into the ER lumen, where they are glycosylated and where molecular chaperones aid protein folding. Misfolded proteins are usually identified here and retrotranslocated by ER-associated degradation to the cytosol, where they are degraded by a proteasome. The vesicles containing the properly folded proteins then enter the Golgi apparatus.
In the Golgi apparatus, the glycosylation of the proteins is modified and further post-translational modifications, including cleavage and functionalization, may occur. The proteins are then moved into secretory vesicles which travel along the cytoskeleton to the edge of the cell. More modification can occur in the secretory vesicles (for example insulin is cleaved from proinsulin in the secretory vesicles).
Eventually, there is vesicle fusion with the cell membrane at porosomes, by a process called exocytosis, dumping its contents out of the cell's environment.
Strict biochemical control is maintained over this sequence by usage of a pH gradient: the pH of the cytosol is 7.4, the ER's pH is 7.0, and the cis-golgi has a pH of 6.5. Secretory vesicles have pHs ranging between 5.0 and 6.0; some secretory vesicles evolve into lysosomes, which have a pH of 4.8.
There are many proteins like FGF1 (aFGF), FGF2 (bFGF), interleukin-1 (IL1) etc. which do not have a signal sequence. They do not use the classical ER-Golgi pathway. These are secreted through various nonclassical pathways.
At least four nonclassical (unconventional) protein secretion pathways have been described. They include:
Secretion
Secretion is the movement of material from one point to another, such as a secreted chemical substance from a cell or gland. In contrast, excretion is the removal of certain substances or waste products from a cell or organism. The classical mechanism of cell secretion is via secretory portals at the plasma membrane called porosomes. Porosomes are permanent cup-shaped lipoprotein structures embedded in the cell membrane, where secretory vesicles transiently dock and fuse to release intra-vesicular contents from the cell.
Secretion in bacterial species means the transport or translocation of effector molecules. For example: proteins, enzymes or toxins (such as cholera toxin in pathogenic bacteria e.g. Vibrio cholerae) from across the interior (cytoplasm or cytosol) of a bacterial cell to its exterior. Secretion is a very important mechanism in bacterial functioning and operation in their natural surrounding environment for adaptation and survival.
Eukaryotic cells, including human cells, have a highly evolved process of secretion. Proteins targeted for the outside are synthesized by ribosomes docked to the rough endoplasmic reticulum (ER). As they are synthesized, these proteins translocate into the ER lumen, where they are glycosylated and where molecular chaperones aid protein folding. Misfolded proteins are usually identified here and retrotranslocated by ER-associated degradation to the cytosol, where they are degraded by a proteasome. The vesicles containing the properly folded proteins then enter the Golgi apparatus.
In the Golgi apparatus, the glycosylation of the proteins is modified and further post-translational modifications, including cleavage and functionalization, may occur. The proteins are then moved into secretory vesicles which travel along the cytoskeleton to the edge of the cell. More modification can occur in the secretory vesicles (for example insulin is cleaved from proinsulin in the secretory vesicles).
Eventually, there is vesicle fusion with the cell membrane at porosomes, by a process called exocytosis, dumping its contents out of the cell's environment.
Strict biochemical control is maintained over this sequence by usage of a pH gradient: the pH of the cytosol is 7.4, the ER's pH is 7.0, and the cis-golgi has a pH of 6.5. Secretory vesicles have pHs ranging between 5.0 and 6.0; some secretory vesicles evolve into lysosomes, which have a pH of 4.8.
There are many proteins like FGF1 (aFGF), FGF2 (bFGF), interleukin-1 (IL1) etc. which do not have a signal sequence. They do not use the classical ER-Golgi pathway. These are secreted through various nonclassical pathways.
At least four nonclassical (unconventional) protein secretion pathways have been described. They include:
Recent media
Recent media