Recent from talks
Rhizaria
Knowledge base stats:
Talk channels stats:
Members stats:
Rhizaria
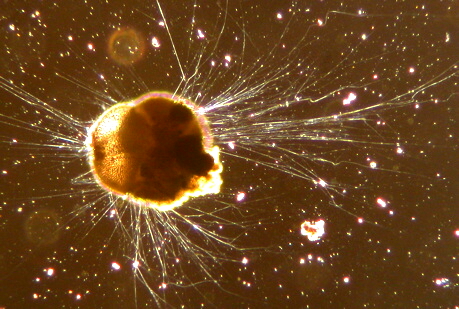
The Rhizaria are a diverse and species-rich clade of mostly unicellular eukaryotes. Except for the chlorarachniophytes and three species in the genus Paulinella in the phylum Cercozoa, they are all non-photosynthetic, but many Foraminifera and Radiolaria have a symbiotic relationship with unicellular algae. A multicellular form, Guttulinopsis vulgaris, a cellular slime mold, has been described. This group was used by Cavalier-Smith in 2002, although the term "Rhizaria" had been long used for clades within the currently recognized taxon.
Being described mainly from rDNA sequences, they vary considerably in form, having no clear morphological distinctive characters (synapomorphies), but for the most part they are amoeboids with filose, reticulose, or microtubule-supported pseudopods. In the absence of an apomorphy, the group is ill-defined, and its composition has been very fluid. Some Rhizaria possess mineral exoskeletons (thecae or loricas), which are in different clades within Rhizaria made out of opal (SiO2), celestite (SrSO4), or calcite (CaCO3).
Certain species can attain sizes of more than a centimeter with some species being able to form cylindrical colonies approximately 1 cm in diameter and greater than 1 m in length. They feed by capturing and engulfing prey with the extensions of their pseudopodia; forms that are symbiotic with unicellular algae contribute significantly to the total primary production of the ocean.
The three main groups of Rhizaria are:
A few other groups may be included in the Cercozoa, but some trees appear closer to the Foraminifera. These are the Phytomyxea and Ascetosporea, parasites of plants and animals, respectively, and the peculiar amoeba Gromia. The different groups of Rhizaria are considered close relatives based mainly on genetic similarities, and have been regarded as an extension of the Cercozoa. The name Rhizaria for the expanded group was introduced by Cavalier-Smith in 2002, who also included the centrohelids and Apusozoa.
A noteworthy order that belongs to Ascetosporea is the Mikrocytida. These are parasites of oysters. This includes the causative agent of Denman Island Disease, Mikrocytos mackini a small (2−3 μm diameter) amitochondriate protistan.
Similarities between various Rhizaria organisms have been noticed since the 19th century. In his 1861 classification of the Rhizopoda (amoebae), the zoologist William B. Carpenter proposed the order Reticularia, which consisted of Foraminifera and Gromiida on the basis of their very similar thin, reticulose pseudopodia with granules circulating inside. However, the idea that these organisms and others such as Radiolaria were all related to one another emerged rather recently, with the help of molecular phylogenetics and advanced microscopy techniques in the late 20th century.
Rhizaria are part of the SAR supergroup (Stramenopiles, Alveolates, Rhizaria), a grouping that had been presaged in 1993 through a study of mitochondrial morphologies. SAR is currently placed in the Diaphoretickes along with Archaeplastida, Cryptista, Haptista, and several minor clades.
Hub AI
Rhizaria AI simulator
(@Rhizaria_simulator)
Rhizaria
The Rhizaria are a diverse and species-rich clade of mostly unicellular eukaryotes. Except for the chlorarachniophytes and three species in the genus Paulinella in the phylum Cercozoa, they are all non-photosynthetic, but many Foraminifera and Radiolaria have a symbiotic relationship with unicellular algae. A multicellular form, Guttulinopsis vulgaris, a cellular slime mold, has been described. This group was used by Cavalier-Smith in 2002, although the term "Rhizaria" had been long used for clades within the currently recognized taxon.
Being described mainly from rDNA sequences, they vary considerably in form, having no clear morphological distinctive characters (synapomorphies), but for the most part they are amoeboids with filose, reticulose, or microtubule-supported pseudopods. In the absence of an apomorphy, the group is ill-defined, and its composition has been very fluid. Some Rhizaria possess mineral exoskeletons (thecae or loricas), which are in different clades within Rhizaria made out of opal (SiO2), celestite (SrSO4), or calcite (CaCO3).
Certain species can attain sizes of more than a centimeter with some species being able to form cylindrical colonies approximately 1 cm in diameter and greater than 1 m in length. They feed by capturing and engulfing prey with the extensions of their pseudopodia; forms that are symbiotic with unicellular algae contribute significantly to the total primary production of the ocean.
The three main groups of Rhizaria are:
A few other groups may be included in the Cercozoa, but some trees appear closer to the Foraminifera. These are the Phytomyxea and Ascetosporea, parasites of plants and animals, respectively, and the peculiar amoeba Gromia. The different groups of Rhizaria are considered close relatives based mainly on genetic similarities, and have been regarded as an extension of the Cercozoa. The name Rhizaria for the expanded group was introduced by Cavalier-Smith in 2002, who also included the centrohelids and Apusozoa.
A noteworthy order that belongs to Ascetosporea is the Mikrocytida. These are parasites of oysters. This includes the causative agent of Denman Island Disease, Mikrocytos mackini a small (2−3 μm diameter) amitochondriate protistan.
Similarities between various Rhizaria organisms have been noticed since the 19th century. In his 1861 classification of the Rhizopoda (amoebae), the zoologist William B. Carpenter proposed the order Reticularia, which consisted of Foraminifera and Gromiida on the basis of their very similar thin, reticulose pseudopodia with granules circulating inside. However, the idea that these organisms and others such as Radiolaria were all related to one another emerged rather recently, with the help of molecular phylogenetics and advanced microscopy techniques in the late 20th century.
Rhizaria are part of the SAR supergroup (Stramenopiles, Alveolates, Rhizaria), a grouping that had been presaged in 1993 through a study of mitochondrial morphologies. SAR is currently placed in the Diaphoretickes along with Archaeplastida, Cryptista, Haptista, and several minor clades.
Recent media