Recent from talks
Vampyrellida
Knowledge base stats:
Talk channels stats:
Members stats:
Vampyrellida
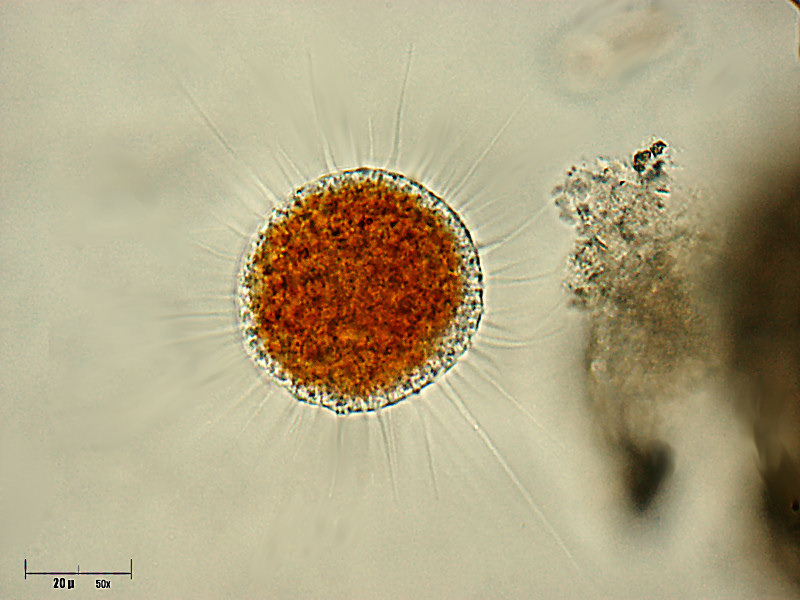
The vampyrellids (order Vampyrellida, class Vampyrellidea), colloquially known as vampire amoebae, are a group of free-living predatory amoebae classified as part of the lineage Endomyxa. They are distinguished from other groups of amoebae by their irregular cell shape with propensity to fuse and split like plasmodial organisms, and their life cycle with a digestive cyst stage that digests the gathered food. They appear worldwide in marine, brackish, freshwater and soil habitats. They are important predators of an enormous variety of microscopic organisms, from algae to fungi and animals. They are also known as aconchulinid amoebae (order Aconchulinida).
Vampyrellids are traditionally considered filose amoebae, i.e. they generate slender pseudopodia (filopodia). They are naked, devoid of external structures such as scales, cell coats or a glycocalyx, although there may be a temporary mucilage coat in the trophozoite stage. The trophozoites vary greatly in shape, size and color between species, but can be grouped into three cell states or 'morphotypes': isodiametric, expanded, and 'filoflabellate'.
All known vampyrellids are heterotrophic amoebae with a free-living (non-parasitic) life cycle that lacks flagellate stages, except for Lateromyxa gallica, and is characterized by an alternation between mobile and immobile cellular stages:
In some species, near the end of the digestive cyst stage, asexual reproduction takes place inside the cyst through a cell division (called 'internal plasmotomy'), resulting in 2–4 daughter cells. These cells are released as young trophozoites through the holes. Other species do not divide inside the closed cyst, and instead divide during or after the hatching process ('external plasmotomy'). Lateromyxa gallica shows an unusual mode of reproduction: while feeding on the inside of algal cells, the plasmodia shed and develop into digestive cysts.
There is a lack of evidence for sexual reproduction in vampyrellids, except for some meiotic stages in resting cysts revealed in Lateromyxa gallica through ultrastructural studies.
Many vampyrellid species have more than one nucleus and behave like plasmodia. They can fuse their cells upon contact, and split apart when moving in opposite directions. Some species readily grow plasmodia as large as a Petri dish under laboratory conditions, while others only fuse when the cell density is high and the food availability is low. It is uncertain to what extend this can happen in the natural environment. In contrast, Placopus species are rarely ever seen with more than two nuclei.
Under adverse environmental conditions, vampyrellids can transform into several types of resting stages:
Vampyrellids have a cosmopolitan distribution: they appear in all continents except Antarctica and all marine ecosystems. They inhabit a wide range of marine, brackish and freshwater habitats, and are frequently isolated from soil samples. Marine ecosystems hold a surprisingly high diversity, and they are found mostly in benthic habitats (e.g. tidal pools, diatom lawns, associated with red algae...). There is a significant positive correlation between the diversity of Vampyrellida and the nutrient availability in the sediment. According to environmental sequencing vampyrellids colonize neotropical soil, glacial cryoconite systems, Brassicaceae leaves, Sphagnum-inhabited peat bogs, hydrothermal sediments and the deep sea.
Hub AI
Vampyrellida AI simulator
(@Vampyrellida_simulator)
Vampyrellida
The vampyrellids (order Vampyrellida, class Vampyrellidea), colloquially known as vampire amoebae, are a group of free-living predatory amoebae classified as part of the lineage Endomyxa. They are distinguished from other groups of amoebae by their irregular cell shape with propensity to fuse and split like plasmodial organisms, and their life cycle with a digestive cyst stage that digests the gathered food. They appear worldwide in marine, brackish, freshwater and soil habitats. They are important predators of an enormous variety of microscopic organisms, from algae to fungi and animals. They are also known as aconchulinid amoebae (order Aconchulinida).
Vampyrellids are traditionally considered filose amoebae, i.e. they generate slender pseudopodia (filopodia). They are naked, devoid of external structures such as scales, cell coats or a glycocalyx, although there may be a temporary mucilage coat in the trophozoite stage. The trophozoites vary greatly in shape, size and color between species, but can be grouped into three cell states or 'morphotypes': isodiametric, expanded, and 'filoflabellate'.
All known vampyrellids are heterotrophic amoebae with a free-living (non-parasitic) life cycle that lacks flagellate stages, except for Lateromyxa gallica, and is characterized by an alternation between mobile and immobile cellular stages:
In some species, near the end of the digestive cyst stage, asexual reproduction takes place inside the cyst through a cell division (called 'internal plasmotomy'), resulting in 2–4 daughter cells. These cells are released as young trophozoites through the holes. Other species do not divide inside the closed cyst, and instead divide during or after the hatching process ('external plasmotomy'). Lateromyxa gallica shows an unusual mode of reproduction: while feeding on the inside of algal cells, the plasmodia shed and develop into digestive cysts.
There is a lack of evidence for sexual reproduction in vampyrellids, except for some meiotic stages in resting cysts revealed in Lateromyxa gallica through ultrastructural studies.
Many vampyrellid species have more than one nucleus and behave like plasmodia. They can fuse their cells upon contact, and split apart when moving in opposite directions. Some species readily grow plasmodia as large as a Petri dish under laboratory conditions, while others only fuse when the cell density is high and the food availability is low. It is uncertain to what extend this can happen in the natural environment. In contrast, Placopus species are rarely ever seen with more than two nuclei.
Under adverse environmental conditions, vampyrellids can transform into several types of resting stages:
Vampyrellids have a cosmopolitan distribution: they appear in all continents except Antarctica and all marine ecosystems. They inhabit a wide range of marine, brackish and freshwater habitats, and are frequently isolated from soil samples. Marine ecosystems hold a surprisingly high diversity, and they are found mostly in benthic habitats (e.g. tidal pools, diatom lawns, associated with red algae...). There is a significant positive correlation between the diversity of Vampyrellida and the nutrient availability in the sediment. According to environmental sequencing vampyrellids colonize neotropical soil, glacial cryoconite systems, Brassicaceae leaves, Sphagnum-inhabited peat bogs, hydrothermal sediments and the deep sea.
Recent media