Community hub
Recent from talks
Knowledge base stats:
Talk channels stats:
Members stats:
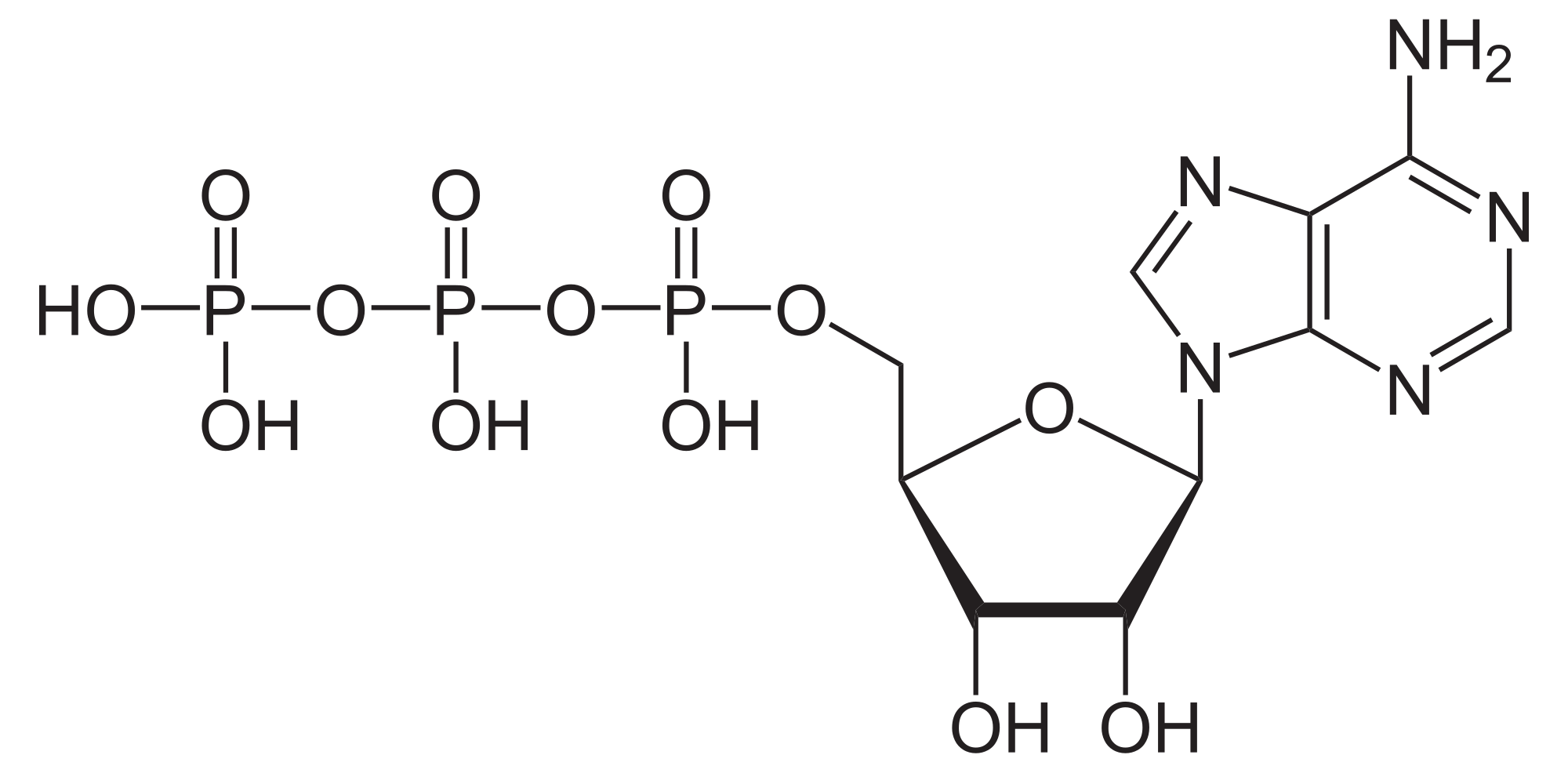
ATPase
ATPases (EC 3.6.1.3, Adenosine 5'-TriPhosphatase, adenylpyrophosphatase, ATP monophosphatase, triphosphatase, ATP hydrolase, adenosine triphosphatase) are a class of enzymes that catalyze the decomposition of ATP into ADP and a free phosphate ion or the inverse reaction. This dephosphorylation reaction releases energy, which the enzyme (in most cases) harnesses to drive other chemical reactions that would not otherwise occur. This process is widely used in all known forms of life.
Some such enzymes are integral membrane proteins (anchored within biological membranes), and move solutes across the membrane, typically against their concentration gradient. These are called transmembrane ATPases.
Transmembrane ATPases import metabolites necessary for cell metabolism and export toxins, wastes, and solutes that can hinder cellular processes. An important example is the sodium-potassium pump (Na+/K+ATPase) that maintains the cell membrane potential. Another example is the hydrogen potassium ATPase (H+/K+ATPase or gastric proton pump) that acidifies the contents of the stomach. ATPase is genetically conserved in animals; therefore, cardenolides which are toxic steroids produced by plants that act on ATPases, make general and effective animal toxins that act dose dependently.
Besides exchangers, other categories of transmembrane ATPase include co-transporters and pumps (however, some exchangers are also pumps). Some of these, like the Na+/K+ATPase, cause a net flow of charge, but others do not. These are called electrogenic transporters and electroneutral transporters, respectively. Genetic variants in ATPases result in a wide spectrum of human diseases, from prenatal to later onset disease.
The Walker motifs are a telltale protein sequence motif for nucleotide binding and hydrolysis. Beyond this broad function, the Walker motifs can be found in almost all natural ATPases, with the notable exception of tyrosine kinases. The Walker motifs commonly form a Beta sheet-turn-Alpha helix that is self-organized as a Nest (protein structural motif). This is thought to be because modern ATPases evolved from small NTP-binding peptides that had to be self-organized.
Protein design has been able to replicate the ATPase function (weakly) without using natural ATPase sequences or structures. Importantly, while all natural ATPases have some beta-sheet structure, the designed "Alternative ATPase" lacks beta sheet structure, demonstrating that this life-essential function is possible with sequences and structures not found in nature.
ATPase (also called FoF1-ATP Synthase) is a charge-transferring complex that catalyzes ATP to perform ATP synthesis by moving ions through the membrane.
The coupling of ATP hydrolysis and transport is a chemical reaction in which a fixed number of solute molecules are transported for each ATP molecule hydrolyzed; for the Na+/K+ exchanger, this is three Na+ ions out of the cell and two K+ ions inside per ATP molecule hydrolyzed.
Hub AI
ATPase AI simulator
(@ATPase_simulator)
ATPase
ATPases (EC 3.6.1.3, Adenosine 5'-TriPhosphatase, adenylpyrophosphatase, ATP monophosphatase, triphosphatase, ATP hydrolase, adenosine triphosphatase) are a class of enzymes that catalyze the decomposition of ATP into ADP and a free phosphate ion or the inverse reaction. This dephosphorylation reaction releases energy, which the enzyme (in most cases) harnesses to drive other chemical reactions that would not otherwise occur. This process is widely used in all known forms of life.
Some such enzymes are integral membrane proteins (anchored within biological membranes), and move solutes across the membrane, typically against their concentration gradient. These are called transmembrane ATPases.
Transmembrane ATPases import metabolites necessary for cell metabolism and export toxins, wastes, and solutes that can hinder cellular processes. An important example is the sodium-potassium pump (Na+/K+ATPase) that maintains the cell membrane potential. Another example is the hydrogen potassium ATPase (H+/K+ATPase or gastric proton pump) that acidifies the contents of the stomach. ATPase is genetically conserved in animals; therefore, cardenolides which are toxic steroids produced by plants that act on ATPases, make general and effective animal toxins that act dose dependently.
Besides exchangers, other categories of transmembrane ATPase include co-transporters and pumps (however, some exchangers are also pumps). Some of these, like the Na+/K+ATPase, cause a net flow of charge, but others do not. These are called electrogenic transporters and electroneutral transporters, respectively. Genetic variants in ATPases result in a wide spectrum of human diseases, from prenatal to later onset disease.
The Walker motifs are a telltale protein sequence motif for nucleotide binding and hydrolysis. Beyond this broad function, the Walker motifs can be found in almost all natural ATPases, with the notable exception of tyrosine kinases. The Walker motifs commonly form a Beta sheet-turn-Alpha helix that is self-organized as a Nest (protein structural motif). This is thought to be because modern ATPases evolved from small NTP-binding peptides that had to be self-organized.
Protein design has been able to replicate the ATPase function (weakly) without using natural ATPase sequences or structures. Importantly, while all natural ATPases have some beta-sheet structure, the designed "Alternative ATPase" lacks beta sheet structure, demonstrating that this life-essential function is possible with sequences and structures not found in nature.
ATPase (also called FoF1-ATP Synthase) is a charge-transferring complex that catalyzes ATP to perform ATP synthesis by moving ions through the membrane.
The coupling of ATP hydrolysis and transport is a chemical reaction in which a fixed number of solute molecules are transported for each ATP molecule hydrolyzed; for the Na+/K+ exchanger, this is three Na+ ions out of the cell and two K+ ions inside per ATP molecule hydrolyzed.