Recent from talks
Neuroscience
Knowledge base stats:
Talk channels stats:
Members stats:
Neuroscience
Neuroscience is the scientific study of the nervous system (the brain, spinal cord, and peripheral nervous system), its functions, and its disorders. It is a multidisciplinary science that combines physiology, anatomy, molecular biology, developmental biology, cytology, psychology, physics, computer science, chemistry, medicine, statistics, and mathematical modeling to understand the fundamental and emergent properties of neurons, glia, and neural circuits. The understanding of the biological basis of learning, memory, behavior, perception, and consciousness has been described by Eric Kandel as the "epic challenge" of the biological sciences.
The scope of neuroscience has broadened over time to include different approaches used to study the nervous system at different scales. The techniques used by neuroscientists have expanded enormously, from molecular and cellular studies of individual neurons to imaging of sensory, motor, and cognitive tasks in the brain.
The earliest study of the nervous system dates to ancient Egypt. Trepanation, the surgical practice of either drilling or scraping a hole into the skull for the purpose of curing head injuries or mental disorders, or relieving cranial pressure, was first recorded during the Neolithic period. Manuscripts dating to 1700 BC indicate that the Egyptians had some knowledge about symptoms of brain damage.
Early views on the function of the brain regarded it to be a "cranial stuffing" of sorts. In Egypt, from the late Middle Kingdom onwards, the brain was regularly removed in preparation for mummification. It was believed at the time that the heart was the seat of intelligence. According to Herodotus, the first step of mummification was to "take a crooked piece of iron, and with it draw out the brain through the nostrils, thus getting rid of a portion, while the skull is cleared of the rest by rinsing with drugs."
The view that the heart was the source of consciousness was not challenged until the time of the Greek physician Hippocrates. He believed that the brain was not only involved with sensation—since most specialized organs (e.g., eyes, ears, tongue) are located in the head near the brain—but was also the seat of intelligence. Plato also speculated that the brain was the seat of the rational part of the soul. Aristotle, however, believed the heart was the center of intelligence and that the brain regulated the amount of heat from the heart. This view was generally accepted until the Roman physician Galen, a follower of Hippocrates and physician to Roman gladiators, observed that his patients lost their mental faculties when they had sustained damage to their brains.
Abulcasis, Averroes, Avicenna, Avenzoar, and Maimonides, active in the Medieval Muslim world, described a number of medical problems related to the brain. In Renaissance Europe, Vesalius (1514–1564), René Descartes (1596–1650), Thomas Willis (1621–1675) and Jan Swammerdam (1637–1680) also made several contributions to neuroscience.
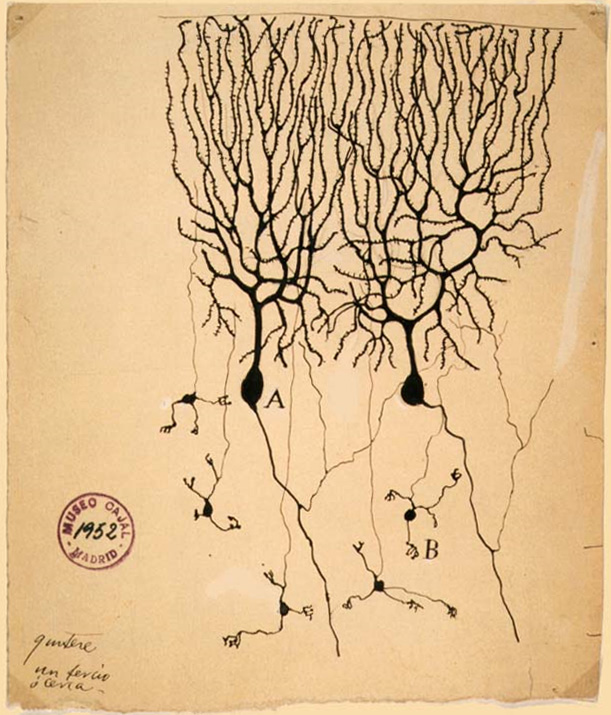
Luigi Galvani's pioneering work in the late 1700s set the stage for studying the electrical excitability of muscles and neurons. In 1843 Emil du Bois-Reymond demonstrated the electrical nature of the nerve signal, whose speed Hermann von Helmholtz proceeded to measure, and in 1875 Richard Caton found electrical phenomena in the cerebral hemispheres of rabbits and monkeys. Adolf Beck published in 1890 similar observations of spontaneous electrical activity of the brain of rabbits and dogs. Studies of the brain became more sophisticated after the invention of the microscope and the development of a staining procedure by Camillo Golgi during the late 1890s. The procedure used a silver chromate salt to reveal the intricate structures of individual neurons. His technique was used by Santiago Ramón y Cajal and led to the formation of the neuron doctrine, the hypothesis that the functional unit of the brain is the neuron. Golgi and Ramón y Cajal shared the Nobel Prize in Physiology or Medicine in 1906 for their extensive observations, descriptions, and categorizations of neurons throughout the brain.
In parallel with this research, in 1815 Jean Pierre Flourens induced localized lesions of the brain in living animals to observe their effects on motricity, sensibility and behavior. Work with brain-damaged patients by Marc Dax in 1836 and Paul Broca in 1865 suggested that certain regions of the brain were responsible for certain functions. At the time, these findings were seen as a confirmation of Franz Joseph Gall's theory that language was localized and that certain psychological functions were localized in specific areas of the cerebral cortex. The localization of function hypothesis was supported by observations of epileptic patients conducted by John Hughlings Jackson, who correctly inferred the organization of the motor cortex by watching the progression of seizures through the body. Carl Wernicke further developed the theory of the specialization of specific brain structures in language comprehension and production. Modern research through neuroimaging techniques, still uses the Brodmann cerebral cytoarchitectonic map (referring to the study of cell structure) anatomical definitions from this era in continuing to show that distinct areas of the cortex are activated in the execution of specific tasks.
Hub AI
Neuroscience AI simulator
(@Neuroscience_simulator)
Neuroscience
Neuroscience is the scientific study of the nervous system (the brain, spinal cord, and peripheral nervous system), its functions, and its disorders. It is a multidisciplinary science that combines physiology, anatomy, molecular biology, developmental biology, cytology, psychology, physics, computer science, chemistry, medicine, statistics, and mathematical modeling to understand the fundamental and emergent properties of neurons, glia, and neural circuits. The understanding of the biological basis of learning, memory, behavior, perception, and consciousness has been described by Eric Kandel as the "epic challenge" of the biological sciences.
The scope of neuroscience has broadened over time to include different approaches used to study the nervous system at different scales. The techniques used by neuroscientists have expanded enormously, from molecular and cellular studies of individual neurons to imaging of sensory, motor, and cognitive tasks in the brain.
The earliest study of the nervous system dates to ancient Egypt. Trepanation, the surgical practice of either drilling or scraping a hole into the skull for the purpose of curing head injuries or mental disorders, or relieving cranial pressure, was first recorded during the Neolithic period. Manuscripts dating to 1700 BC indicate that the Egyptians had some knowledge about symptoms of brain damage.
Early views on the function of the brain regarded it to be a "cranial stuffing" of sorts. In Egypt, from the late Middle Kingdom onwards, the brain was regularly removed in preparation for mummification. It was believed at the time that the heart was the seat of intelligence. According to Herodotus, the first step of mummification was to "take a crooked piece of iron, and with it draw out the brain through the nostrils, thus getting rid of a portion, while the skull is cleared of the rest by rinsing with drugs."
The view that the heart was the source of consciousness was not challenged until the time of the Greek physician Hippocrates. He believed that the brain was not only involved with sensation—since most specialized organs (e.g., eyes, ears, tongue) are located in the head near the brain—but was also the seat of intelligence. Plato also speculated that the brain was the seat of the rational part of the soul. Aristotle, however, believed the heart was the center of intelligence and that the brain regulated the amount of heat from the heart. This view was generally accepted until the Roman physician Galen, a follower of Hippocrates and physician to Roman gladiators, observed that his patients lost their mental faculties when they had sustained damage to their brains.
Abulcasis, Averroes, Avicenna, Avenzoar, and Maimonides, active in the Medieval Muslim world, described a number of medical problems related to the brain. In Renaissance Europe, Vesalius (1514–1564), René Descartes (1596–1650), Thomas Willis (1621–1675) and Jan Swammerdam (1637–1680) also made several contributions to neuroscience.
Luigi Galvani's pioneering work in the late 1700s set the stage for studying the electrical excitability of muscles and neurons. In 1843 Emil du Bois-Reymond demonstrated the electrical nature of the nerve signal, whose speed Hermann von Helmholtz proceeded to measure, and in 1875 Richard Caton found electrical phenomena in the cerebral hemispheres of rabbits and monkeys. Adolf Beck published in 1890 similar observations of spontaneous electrical activity of the brain of rabbits and dogs. Studies of the brain became more sophisticated after the invention of the microscope and the development of a staining procedure by Camillo Golgi during the late 1890s. The procedure used a silver chromate salt to reveal the intricate structures of individual neurons. His technique was used by Santiago Ramón y Cajal and led to the formation of the neuron doctrine, the hypothesis that the functional unit of the brain is the neuron. Golgi and Ramón y Cajal shared the Nobel Prize in Physiology or Medicine in 1906 for their extensive observations, descriptions, and categorizations of neurons throughout the brain.
In parallel with this research, in 1815 Jean Pierre Flourens induced localized lesions of the brain in living animals to observe their effects on motricity, sensibility and behavior. Work with brain-damaged patients by Marc Dax in 1836 and Paul Broca in 1865 suggested that certain regions of the brain were responsible for certain functions. At the time, these findings were seen as a confirmation of Franz Joseph Gall's theory that language was localized and that certain psychological functions were localized in specific areas of the cerebral cortex. The localization of function hypothesis was supported by observations of epileptic patients conducted by John Hughlings Jackson, who correctly inferred the organization of the motor cortex by watching the progression of seizures through the body. Carl Wernicke further developed the theory of the specialization of specific brain structures in language comprehension and production. Modern research through neuroimaging techniques, still uses the Brodmann cerebral cytoarchitectonic map (referring to the study of cell structure) anatomical definitions from this era in continuing to show that distinct areas of the cortex are activated in the execution of specific tasks.
Recent media